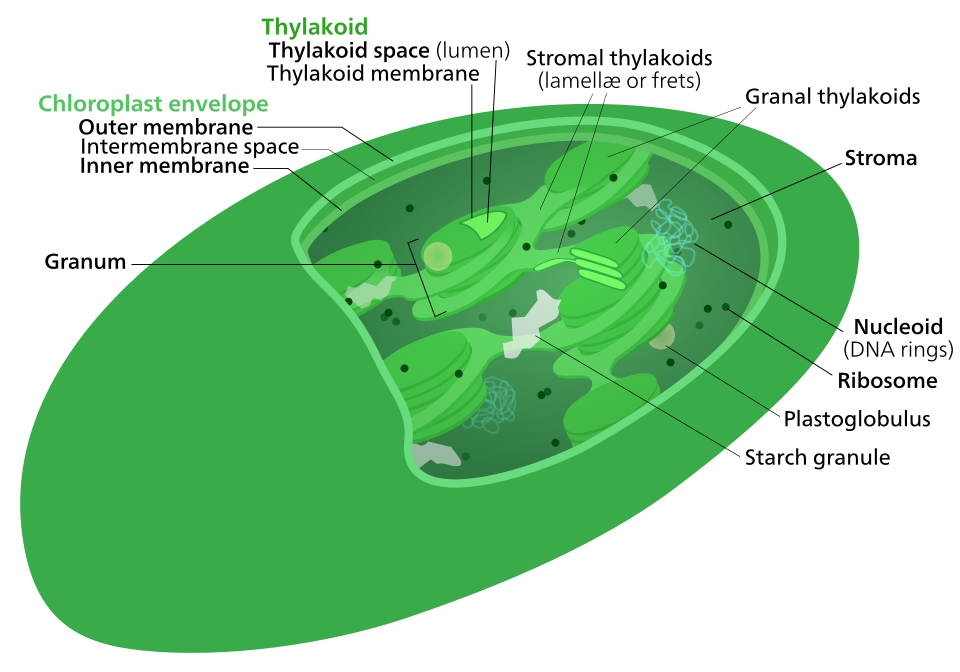
Le cycle de Calvin , également appelé phase sombre de la photosynthèse , est une série de réactions chimiques transformant carbone et les transporteurs d'hydrogène en glucose . Ce cycle est présent chez tous les eucaryotes photosynthétiques et chez de nombreuses bactéries photosynthétiques. Chez les plantes, ces réactions se déroulent dans le stroma la région fluide du chloroplaste située à l'extérieur des membranes thylakoïdes . Elles utilisent les produits ( ATP et NADPH ) des réactions photochimiques pour effectuer d'autres transformations. Le cycle de Calvin exploite l'énergie chimique de l'ATP et le pouvoir réducteur du NADPH issus des réactions photochimiques pour produire des sucres utilisables par la plante. Ces substrats sont utilisés dans une série de réactions d'oxydoréduction ( redox ) pour produire des sucres par étapes ; il n'existe pas de réaction directe convertissant plusieurs molécules de CO₂ en un sucre. Les réactions indépendantes de la lumière, appelées collectivement cycle de Calvin, comportent trois phases : la carboxylation , les réactions de réduction et la régénération du ribulose 1,5-bisphosphate (RuBP).
Bien qu'on l'appelle aussi « phase sombre », le cycle de Calvin ne se déroule pas dans l'obscurité ni la nuit. En effet, ce processus nécessite du NADPH , une molécule éphémère issue des réactions photochimiques . Dans l'obscurité, les plantes libèrent du saccharose dans le phloème à partir de leurs réserves d'amidon pour fournir l'énergie nécessaire à leur croissance. Le cycle de Calvin a donc lieu lorsque la lumière est disponible, indépendamment du type de photosynthèse ( fixation du carbone en C3 , fixation du carbone en C4 et métabolisme acide crassulacéen (CAM) ). Les plantes CAM stockent l'acide malique dans leurs vacuoles chaque nuit et le libèrent le jour pour permettre ce processus.
Couplage à d'autres voies métaboliques
Les réactions du cycle de Calvin sont étroitement liées à la chaîne de transport d'électrons des thylakoïdes , car l'énergie nécessaire à la réduction du dioxyde de carbone est fournie par le NADPH produit lors des réactions photochimiques . La photorespiration , également appelée cycle C2, est aussi liée au cycle de Calvin, car elle résulte d'une réaction alternative de l' enzyme RuBisCO , et son produit final est une autre molécule de glycéraldéhyde-3-phosphate .
cycle Calvin
Le cycle de Calvin , également appelé cycle de Calvin-Benson-Bassham (CBB) , cycle des pentoses phosphates réducteurs (cycle RPP) ou cycle C3, est une série de réactions d'oxydoréduction biochimiques qui se déroulent dans le stroma des chloroplastes des organismes photosynthétiques . Ce cycle a été découvert en 1950 par Melvin Calvin , James Bassham et Andrew Benson à l' Université de Californie à Berkeley, grâce à l' isotope radioactif carbone-14 .
La photosynthèse se déroule en deux étapes dans la cellule. Lors de la première étape, les réactions photochimiques captent l'énergie lumineuse et l'utilisent pour synthétiser l' ATP, molécule de stockage d'énergie, et le NADPH, transporteur d'hydrogène à énergie modérée . Le cycle de Calvin utilise ces composés pour convertir le dioxyde de carbone et l'eau en composés organiques utilisables par l'organisme (et par les animaux qui s'en nourrissent). Cet ensemble de réactions est également appelé fixation du carbone . L' enzyme clé de ce cycle est la RuBisCO . Dans les équations biochimiques suivantes, les espèces chimiques (phosphates et acides carboxyliques) sont en équilibre entre leurs différents états d'ionisation, sous l'influence du pH .
Les enzymes du cycle de Calvin sont fonctionnellement équivalentes à la plupart des enzymes impliquées dans d'autres voies métaboliques, telles que la gluconéogenèse et la voie des pentoses phosphates . Cependant, elles se trouvent dans le stroma des chloroplastes et non dans le cytosol cellulaire , ce qui isole les réactions. Elles sont activées par la lumière (d'où le caractère trompeur de l'appellation « phase sombre ») et par les produits de la phase photochimique. Ces mécanismes de régulation empêchent la respiration cellulaire et la production de dioxyde de carbone. En effet, l'énergie (sous forme d'ATP) serait gaspillée pour ces réactions si leur productivité nette était nulle .
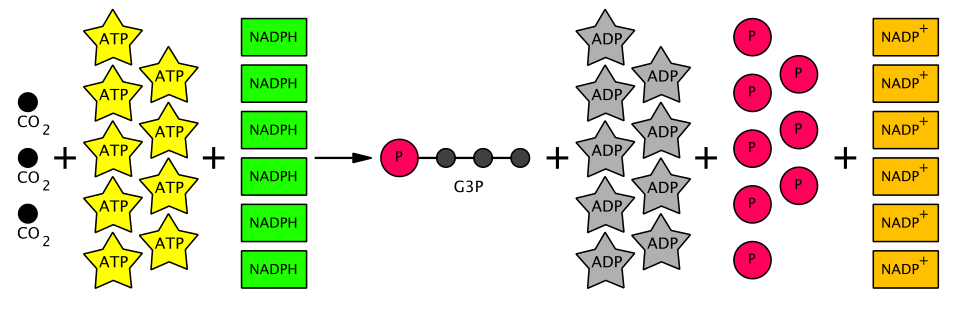
La somme des réactions du cycle de Calvin est la suivante :
- 3 CO2+ 6 NADPH + 9 ATP + 6 H + → glycéraldéhyde-3-phosphate (G3P) + 6 NADP + + 9 ADP + 3 H 2 O + 8 Pi ( Pi = phosphate inorganique )
Les hexoses (sucres à six carbones) ne sont pas des produits du cycle de Calvin. Bien que de nombreux ouvrages mentionnent un produit de la photosynthèse comme étant le C6H12O6Ceci est principalement dû à une volonté de simplification, afin de correspondre à l'équation de la respiration aérobie , où les sucres à six carbones sont oxydés dans les mitochondries. Les produits glucidiques du cycle de Calvin sont des molécules de sucre phosphate à trois carbones, ou « triose phosphates », à savoir le glycéraldéhyde-3-phosphate (G3P).
Mesures
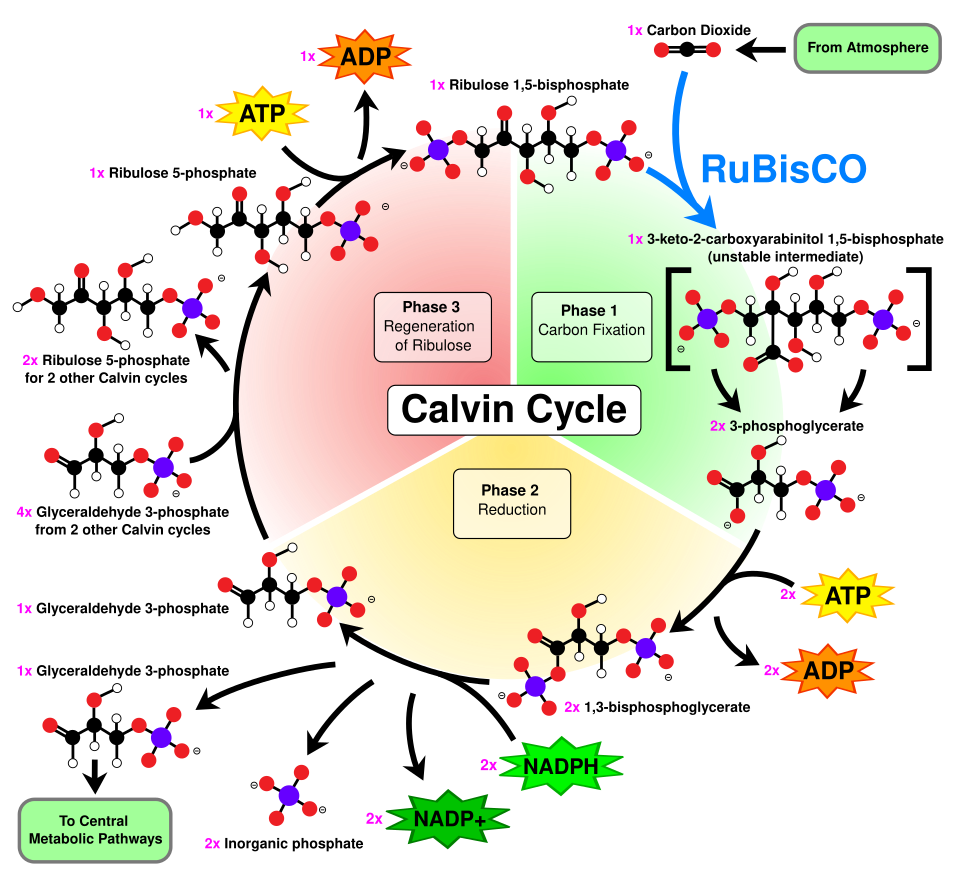
Dans la première étape du cycle de Calvin, une molécule de CO₂ est incorporée à l'une des deux molécules à trois carbones ( glycéraldéhyde-3-phosphate ou G3P), consommant ainsi deux molécules d' ATP et deux molécules de NADPH , produites lors de la phase photochimique. Les trois étapes impliquées sont :
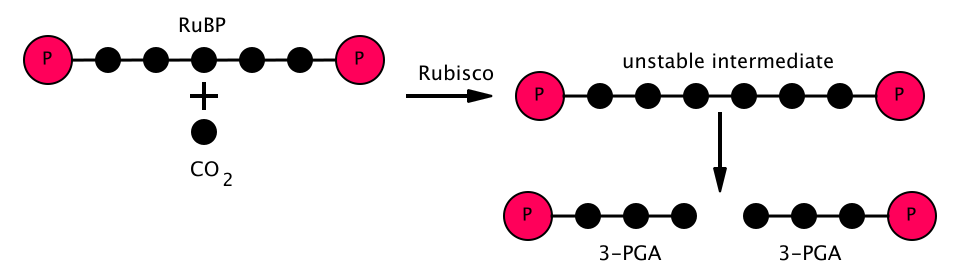

- L'enzyme RuBisCO catalyse la carboxylation du ribulose-1,5-bisphosphate (RuBP), un composé à 5 carbones, par le dioxyde de carbone (6 carbones au total) en deux étapes. Le produit de la première étape est un complexe énédiol-enzyme capable de capter le CO₂.2ou O2Ainsi, le complexe énédiol-enzyme est la véritable carboxylase/oxygénase. Le CO2qui est capturé par l'énédiol dans la deuxième étape produit un composé instable à six carbones appelé 2-carboxy 3-céto 1,5-biphosphoribotol (CKABP ) (ou 3-céto-2-carboxyarabinitol 1,5-bisphosphate) qui se divise immédiatement en 2 molécules de 3-phosphoglycérate (également écrit acide 3-phosphoglycérique, PGA, 3PGA ou 3-PGA), un composé à 3 carbones.
- L'enzyme phosphoglycérate kinase catalyse la phosphorylation du 3-PGA par l'ATP (produit lors de la phase photochimique). Les produits de cette phosphorylation sont le 1,3-bisphosphoglycérate (glycérate-1,3-bisphosphate) et l'ADP . (Il est à noter que deux molécules de 3-PGA sont produites pour chaque molécule de CO₂. )2qui entre dans le cycle, donc cette étape utilise deux ATP par CO22fixé.)
- L'enzyme glycéraldéhyde-3-phosphate déshydrogénase catalyse la réduction du 1,3BPGA par le NADPH (un autre produit de la phase photochimique). Le glycéraldéhyde-3-phosphate (également appelé G3P, GP, TP, PGAL, GAP) est produit, et le NADPH est oxydé en NADP + . Ainsi, deux molécules de NADPH sont consommées par molécule de CO2 .2fixé.
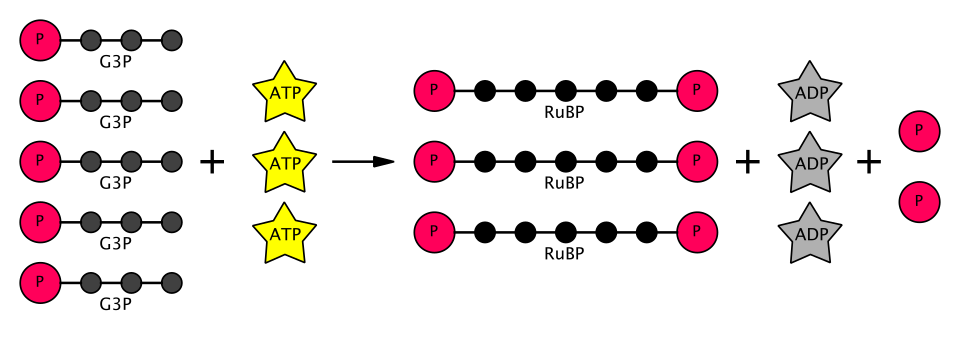
L'étape suivante du cycle de Calvin consiste à régénérer le RuBP . Cinq molécules de G3P produisent trois molécules de RuBP , consommant ainsi trois molécules d'ATP. Puisque chaque molécule de CO₂ produit deux molécules de G3P, trois molécules de CO₂ produisent six molécules de G3P, dont cinq sont utilisées pour régénérer le RuBP , ce qui représente un gain net d'une molécule de G3P pour trois molécules de CO₂ ( comme on pouvait s'y attendre compte tenu du nombre d'atomes de carbone impliqués).


La phase de régénération peut être décomposée en une série d'étapes.
- La triose phosphate isomérase convertit l'un des G3P de manière réversible en dihydroxyacétone phosphate (DHAP), également une molécule à 3 carbones.
- L'aldolase et la fructose-1,6-bisphosphatase transforment un G3P et un DHAP en fructose 6-phosphate (6C). Un ion phosphate est libéré dans la solution.
- Puis fixation d'un autre CO2génère deux G3P supplémentaires.
- La F6P subit l'élimination de deux atomes de carbone par la transkétolase , donnant l'érythrose-4-phosphate (E4P). Ces deux atomes de carbone sont ensuite ajoutés à une G3P, formant le cétose xylulose -5-phosphate (Xu5P).
- E4P et un DHAP (formé à partir d'un des G3P du deuxième CO2fixation) sont convertis en sédoheptulose-1,7-bisphosphate (7C) par l'enzyme aldolase.
- La sédoheptulose-1,7-bisphosphatase (l'une des trois seules enzymes du cycle de Calvin propres aux plantes) clive le sédoheptulose-1,7-bisphosphate en sédoheptulose-7-phosphate , libérant un ion phosphate inorganique en solution.
- Fixation d'un troisième CO2Deux autres molécules de G3P sont générées. Le cétose S7P subit l'élimination de deux carbones par la transkétolase , donnant du ribose-5-phosphate (R5P), et les deux carbones restants sur la transkétolase sont transférés à l'une des molécules de G3P, donnant un autre Xu5P. Il reste ainsi une molécule de G3P comme produit de la fixation de 3 CO₂.2, avec la génération de trois pentoses qui peuvent être convertis en Ru5P.
- Le R5P est converti en ribulose-5-phosphate (Ru5P, RuP) par la phosphopentose isomérase . Le Xu5P est converti en RuP par la phosphopentose épimérase .
- Enfin, la phosphoribulokinase (une autre enzyme spécifique aux plantes de cette voie métabolique) phosphoryle le RuP en RuBP (ribulose-1,5-bisphosphate), achevant ainsi le cycle de Calvin . Cette réaction nécessite l'apport d'une molécule d'ATP.
Ainsi, sur les six G3P produits, cinq sont utilisés pour synthétiser trois molécules de RuBP (5C) (soit 15 carbones au total), une seule molécule de G3P étant disponible pour la conversion ultérieure en hexose. Cette réaction nécessite neuf molécules d'ATP et six molécules de NADPH pour trois molécules de CO₂.2molécules. L'équation du cycle de Calvin global est représentée schématiquement ci-dessous.
La RuBisCO réagit également de manière compétitive avec O2au lieu de CO2Lors de la photorespiration , le taux de photorespiration est plus élevé à haute température. La photorespiration transforme le RuBP en 3-PGA et en 2-phosphoglycolate, une molécule à deux carbones qui peut être convertie en glycine via le glycolate et le glyoxylate. Par le système de clivage de la glycine et le tétrahydrofolate, deux glycines sont converties en sérine et CO₂.2La sérine peut être reconvertie en 3-phosphoglycérate. Ainsi, seuls 3 des 4 carbones de deux phosphoglycolates peuvent être reconvertis en 3-PGA. On constate que la photorespiration a des conséquences très négatives pour la plante, car, au lieu de fixer le CO₂, elle le gaspille.2Ce processus entraîne une perte de CO22La fixation du carbone C4 a évolué pour contourner la photorespiration, mais elle ne peut se produire que chez certaines plantes originaires de climats très chauds ou tropicaux, comme le maïs. De plus, les RuBisCO catalysant les réactions de la phase sombre de la photosynthèse présentent généralement une spécificité accrue pour le CO₂ par rapport à l'O₂ , afin de minimiser la réaction d'oxygénation. Cette spécificité accrue est apparue après l'incorporation d'une nouvelle sous-unité protéique par la RuBisCO.
Produits
Les produits immédiats d'un tour du cycle de Calvin sont deux molécules de glycéraldéhyde-3-phosphate (G3P) ; 3 ADP et 2 NADP + sont également régénérés et réutilisés ultérieurement dans les réactions photochimiques . Chaque molécule de G3P est composée de 3 atomes de carbone. Pour que le cycle de Calvin se poursuive, le RuBP (ribulose-1,5-bisphosphate) doit être régénéré ; 5 des 6 atomes de carbone des 2 molécules de G3P sont utilisés à cette fin. Par conséquent, un seul atome de carbone est produit à chaque tour. La synthèse d'une molécule de G3P en surplus nécessite 3 atomes de carbone, et donc 3 tours du cycle de Calvin. La synthèse d'une molécule de glucose (qui peut être obtenue à partir de 2 molécules de G3P) nécessiterait 6 tours du cycle de Calvin. Le G3P en surplus peut également être utilisé pour former d'autres glucides tels que l'amidon, le saccharose et la cellulose, selon les besoins de la plante.
Régulation dépendante de la lumière
Ces réactions ne se produisent pas dans l'obscurité ni la nuit. Il existe une régulation des enzymes du cycle dépendante de la lumière, car la troisième étape nécessite du NADPH.
Deux systèmes de régulation sont à l'œuvre lorsque le cycle doit être activé ou désactivé : le système d'activation thioredoxine / ferredoxine , qui active certaines enzymes du cycle ; et l' activation de l'enzyme RuBisCo , active dans le cycle de Calvin, qui implique sa propre activase.
Le système thiorédoxine/ferrédoxine active les enzymes glycéraldéhyde-3-phosphate déshydrogénase, glycéraldéhyde-3-phosphate phosphatase, fructose-1,6-bisphosphatase, sédoheptulose-1,7-bisphosphatase et ribulose-5-phosphatase kinase, étapes clés du processus. Cette activation a lieu en présence de lumière, lorsque la ferrédoxine est réduite au sein du photosystème I de la chaîne respiratoire thylakoïde, lors de la circulation des électrons. La ferrédoxine se lie alors à la thiorédoxine et la réduit, ce qui active les enzymes du cycle en rompant une liaison cystéine présente dans toutes ces enzymes. Ce processus est dynamique, car cette même liaison est reformée par d'autres protéines qui désactivent les enzymes. Ainsi, les enzymes restent majoritairement actives le jour et sont désactivées à l'obscurité, en l'absence de ferrédoxine réduite.
L'enzyme RuBisCo possède son propre processus d'activation, plus complexe. Son activation nécessite la carbamylation d'un acide aminé spécifique, la lysine . Cette lysine se lie au RuBP et, si elle n'est pas carbamylée, l'enzyme est inactive. Une enzyme activase spécifique, appelée activase de la RuBisCo , facilite cette carbamylation en déprotonant la lysine et en permettant la fixation du dioxyde de carbone. Même ainsi, l'enzyme RuBisCo n'est pas encore fonctionnelle, car elle a besoin d'un ion magnésium lié à la lysine pour fonctionner. Cet ion magnésium est libéré du lumen thylakoïde lorsque le pH interne diminue sous l'effet du pompage actif de protons par le flux d'électrons. L'activase de la RuBisCo est elle-même activée par l'augmentation de la concentration d'ATP dans le stroma, induite par sa phosphorylation .