Les générateurs de schémas centraux ( GSC ) sont des circuits neuronaux biologiques auto-organisés qui produisent des sorties rythmiques en l'absence d'entrées rythmiques. Ils sont la source des schémas d'activité neuronale étroitement couplés qui entraînent des comportements moteurs rythmiques et stéréotypés comme la marche, la natation, la respiration ou la mastication. La capacité de fonctionner sans apport des zones cérébrales supérieures nécessite toujours des entrées modulatrices , et leurs sorties ne sont pas fixes. La flexibilité en réponse aux entrées sensorielles est une qualité fondamentale du comportement piloté par les GSC. Pour être classé comme générateur rythmique, un GSC nécessite :
- « deux ou plusieurs processus qui interagissent de telle sorte que chaque processus augmente et diminue séquentiellement, et
- que, du fait de cette interaction, le système revient à plusieurs reprises à son état de départ. »
Les CPG sont présents chez les humains et la plupart des autres vertébrés, ainsi que chez certains invertébrés.
Physiologie
Neurones CPG

Les neurones CPG peuvent avoir différentes propriétés membranaires intrinsèques (voir schéma). Certains neurones déclenchent des salves de potentiels d'action, soit de manière endogène, soit en présence de substances neuromodulatrices. D'autres neurones sont bistables et génèrent des potentiels de plateau qui peuvent être déclenchés par une impulsion de courant dépolarisante et terminés par une impulsion de courant hyperpolarisante. De nombreux neurones CPG déclenchent après avoir été libérés de l'inhibition (rebond post-inhibiteur). Une autre caractéristique commune des neurones CPG est une diminution de la fréquence de déclenchement lors d'une dépolarisation constante (adaptation de la fréquence des pics).
Génération de rythme

La génération du rythme dans les réseaux CPG dépend des propriétés intrinsèques des neurones CPG et de leurs connexions synaptiques. Il existe deux mécanismes généraux de génération du rythme : le stimulateur/suiveur et l'inhibition réciproque (voir schéma).
Dans un réseau piloté par un stimulateur cardiaque, un ou plusieurs neurones agissent comme un oscillateur central (stimulateur cardiaque) qui entraîne d'autres neurones non pulsés (suiveurs) dans un modèle rythmique. Parmi les exemples de réseaux pilotés par un stimulateur cardiaque, on peut citer le rythme pylorique du ganglion stomatogastrique des crustacés et les rythmes respiratoires des vertébrés.
Dans un réseau piloté par inhibition réciproque, deux (groupes de) neurones s'inhibent mutuellement. De tels réseaux sont connus sous le nom d'oscillateurs à demi-centre. Les neurones ne sont pas actifs rythmiquement lorsqu'ils sont isolés, mais ils peuvent produire des schémas d'activité alternés lorsqu'ils sont couplés par des connexions inhibitrices. (Les neurones peuvent également produire des schémas d'activité d'autres phases relatives, y compris la synchronie, en fonction des propriétés synaptiques). Les transitions entre les états activés et inhibés peuvent se produire via un certain nombre de mécanismes. Par exemple, l'adaptation de la fréquence des pics dans le ou les neurones en rafale peut lentement libérer le ou les autres neurones de l'inhibition. L'inhibition réciproque est une caractéristique essentielle de nombreux CPG, y compris ceux impliqués dans la locomotion.
Les jonctions communicantes contribuent également aux oscillations rythmiques et à la synchronie neuronale dans les CPG. Elles agissent comme un filtre passe-bas permettant aux fluctuations lentes de la tension membranaire de passer plus efficacement à travers les cellules. Chez les souris néonatales, le blocage des jonctions communicantes entraîne une diminution de l'activité rythmique et peut complètement abolir la locomotion fictive induite par le médicament. Chez le poisson zèbre, les motoneurones contrôlent rétrogradement la fréquence de nage via les jonctions communicantes.
Dynamique synaptique à court terme
Les réseaux CPG présentent de nombreuses connexions synaptiques récurrentes, notamment une excitation et une inhibition réciproques. Les synapses des réseaux CPG sont soumises à des modifications dépendantes de l'activité à court terme. La dépression synaptique à court terme et la facilitation des synapses peuvent jouer un rôle dans les transitions entre les phases actives et inactives de l'éclatement et la fin des éclatements.
Circuits CPG
Les circuits CPG supposés être impliqués dans le contrôle de la locomotion sont constitués de motoneurones et d'interneurones spinaux et sont situés dans les régions thoraciques et lombaires inférieures de la moelle épinière des vertébrés , et dans chaque neuromère de la moelle épinière ventrale des invertébrés . Les neurones CPG impliqués dans la déglutition sont situés dans le tronc cérébral, plus précisément dans le noyau hypoglosse de la moelle épinière.
Bien que l'emplacement général des neurones CPG puisse souvent être déduit, l'emplacement spécifique et l'identité des neurones participants commencent seulement à être compris. Les réseaux CPG sont souvent distribués et peuvent être réorganisés de manière flexible, ce qui rend difficile l'identification des interneurones spinaux. Au cours des dernières décennies, des programmes moléculaires et génétiques qui contrôlent la configuration neuronale ont été utilisés pour cibler spécifiquement les interneurones spinaux chez la souris et le poisson zèbre. Le tube neural en développement de la souris embryonnaire montre l'expression de facteurs de transcription distincts dans des domaines le long de l'axe dorso-ventral de la moelle épinière. Ces domaines donnent naissance à une population distincte de neurones qui ont été classés comme classes cardinales dorsales (dI1-dI6) et ventrales (V0-V3) d'interneurones spinaux. Les neurones ventraux sont considérés comme des membres du réseau CPG spinal. Chacune de ces classes d'interneurones peut être divisée en diverses sous-populations de neurones avec un phénotype de neurotransmetteur, une projection axonale et une fonction distinctes pendant la locomotion. Par exemple, les interneurones V2 sont à projection ipsilatérale et peuvent être classés en V2a excitateurs et V2b inhibiteurs. Les V2 sont importants pour l'alternance des fléchisseurs et des extenseurs et fournissent une excitation aux motoneurones.
Neuromodulation
Les organismes doivent adapter leur comportement pour répondre aux besoins de leur environnement interne et externe. Les générateurs de schémas centraux, en tant que partie du circuit neuronal d'un organisme, peuvent être modulés pour s'adapter aux besoins et à l'environnement de l'organisme. Trois rôles de neuromodulation ont été identifiés pour les circuits CPG :
- La modulation est intrinsèque au réseau CPG ou nécessaire à son activation
- La modulation modifie la configuration fonctionnelle des CPG pour produire des sorties différentes
- La modulation modifie le complément des neurones CPG en commutant les neurones entre les réseaux et en fusionnant des réseaux autrefois séparés en entités plus grandes
Les synapses neuromodulatrices peuvent faire partie du réseau CPG lui-même. Par exemple, le CPG sous-jacent à la réponse de nage d'échappement chez Tritonia diomedea contient des neurones neuromodulateurs intrinsèques. Ces neurones neuromodulateurs peuvent améliorer la libération de neurotransmetteurs par un autre neurone du circuit et ses actions neuromodulatrices sont considérées comme importantes pour produire un programme moteur de nage. Les entrées neuromodulatrices peuvent également activer les réseaux CPG et peuvent être nécessaires à la génération d'une sortie rythmique. La perte d'entrées neuromodulatrices peut abolir l'activité rythmique du réseau pylorique. Chez les vertébrés, il a été démontré que l'application de neuromodulateurs évoque une activité locomotrice.
Les neuromodulateurs peuvent modifier la force synaptique ainsi que les propriétés intrinsèques des neurones. Ces actions peuvent modifier la fréquence et les relations de phase entre les neurones et ainsi modifier le modèle de sortie du circuit. Par exemple, l'application exogène de divers neuromodulateurs peut provoquer un modèle moteur triphasique dans le STG où chaque modulateur entraîne la génération d'un modèle moteur différent. Les projections neuromodulatrices exprimant un modulateur commun peuvent également provoquer des modèles différents à partir du même réseau. La stimulation de différents neurones de projection contenant de la proctoline dans le STG entraîne un modèle moteur distinct à partir du même réseau en raison des différences dans le complément de cotransmetteurs de ces neurones de projection.
Les effets des neuromodulateurs sont répartis dans tout le réseau CPG. En particulier, il a été démontré que la dopamine affecte les propriétés cellulaires et synaptiques de presque tous les composants du réseau pylorique des crustacés. De plus, la dopamine peut avoir des effets opposés sur différents composants du réseau. Ainsi, le résultat final du réseau reflète une combinaison d'actions modulatrices sur des composants individuels.
La modulation altère les neurones CPG
La composition neuronale des CPG peut varier en fonction de l'état du système. Les neuromodulateurs peuvent activer ou inhiber les neurones des CPG et peuvent même combiner différents réseaux en un seul. Par exemple, dans le système nerveux stomatogastrique du homard, le neuropeptide, l'hormone de concentration du pigment rouge, peut renforcer les synapses entre deux réseaux différents pour créer un rythme unique et combiné. Les neuromodulateurs peuvent également entraîner le passage de neurones d'un réseau à un autre.
Rétroaction sensorielle
Bien que la théorie de la génération de schémas centraux exige que la rythmicité et le schéma de base soient générés de manière centrale, les CPG peuvent répondre à la rétroaction sensorielle pour modifier le schéma de manière comportementale appropriée. La modification du schéma est difficile car la rétroaction reçue pendant une seule phase peut nécessiter un mouvement modifié dans les autres parties du cycle structuré pour préserver certaines relations de coordination. Par exemple, marcher avec un caillou dans la chaussure droite modifie toute la démarche, même si le stimulus n'est présent que lorsque l'on se tient debout sur le pied droit. Même pendant le temps où le pied gauche est au sol et que la rétroaction sensorielle est inactive, des mesures sont prises pour prolonger le balancement de la jambe droite et prolonger le temps sur le pied gauche, ce qui conduit à boiter. Cet effet pourrait être dû aux effets étendus et durables de la rétroaction sensorielle sur le CPG ou aux effets à court terme sur quelques neurones qui à leur tour modulent les neurones voisins et propagent la rétroaction à travers l'ensemble du CPG de cette manière. Un certain degré de modulation est nécessaire pour permettre à un CPG d'assumer plusieurs états en réponse à la rétroaction.
De plus, l'effet de l'entrée sensorielle varie en fonction de la phase du schéma dans laquelle elle se produit. Par exemple, pendant la marche, la résistance exercée sur le haut du pied oscillant (par exemple, par un bâton horizontal) fait que le pied se lève plus haut pour se déplacer sur le bâton. Cependant, la même entrée sur le pied d'appui ne peut pas faire lever le pied, sinon la personne s'effondrerait. Ainsi, selon la phase, la même entrée sensorielle peut faire lever le pied plus haut ou le maintenir plus fermement au sol. « Ce changement de réponse motrice en fonction de la phase du schéma moteur est appelé inversion réflexe et a été observé chez les invertébrés (DiCaprio et Clarac, 1981) et les vertébrés (Forssberg et al., 1977). La manière dont ce processus se produit est mal comprise, mais là encore deux possibilités existent. L'une est que l'entrée sensorielle est correctement acheminée vers différents neurones CPG en fonction de la phase du schéma moteur. L'autre est que l'entrée atteint les mêmes neurones à toutes les phases, mais que, en conséquence de la manière dont le réseau transforme l'entrée, la réponse du réseau varie de manière appropriée en fonction de la phase du schéma moteur. »
Une étude réalisée par Gottschall et Nichols a examiné le membre postérieur d'un chat décérébré pendant la marche (une fonction contrôlée par le CPG) en réponse aux changements de l'inclinaison de la tête. Cette étude décrit les différences de démarche et de position du corps des chats marchant en montée, en descente et sur des surfaces planes. Les récepteurs proprioceptifs (organes tendineux de Golgi et fuseaux musculaires) et extéroceptifs (optiques, vestibulaires et cutanés) fonctionnent seuls ou en combinaison pour ajuster le CPG au retour sensoriel. L'étude a exploré les effets des propriocepteurs du cou (donnant des informations sur la position relative de la tête et du corps) et des récepteurs vestibulaires (donnant des informations sur l'orientation de la tête par rapport à la gravité). Les chats décérébrés ont été obligés de marcher sur une surface plane avec la tête à niveau, inclinée vers le haut ou vers le bas. La comparaison des chats décérébrés avec des chats normaux a montré des schémas EMG similaires pendant la marche à niveau et des schémas EMG qui reflétaient la marche en descente avec la tête inclinée vers le haut et la marche en montée avec la tête inclinée vers le bas. Cette étude a démontré que les propriocepteurs du cou et les récepteurs vestibulaires contribuent à la rétroaction sensorielle qui modifie la démarche de l'animal. Ces informations peuvent être utiles pour le traitement des troubles de la marche.
Fonctions
Les générateurs de motifs centraux peuvent remplir de nombreuses fonctions. Les CPG peuvent jouer un rôle dans le mouvement, la respiration, la génération de rythme et d'autres fonctions oscillatoires . Vous trouverez ci-dessous plusieurs fonctions clés des CPG.
Locomotion
Dès 1911, les expériences de Thomas Graham Brown ont montré que le modèle de base de la marche peut être produit par la moelle épinière sans qu'il soit nécessaire de recevoir des commandes descendantes du cortex.
La première preuve moderne de l'existence d'un générateur de motifs centraux a été apportée en isolant le système nerveux du criquet et en montrant qu'il pouvait produire de manière isolée une sortie rythmique ressemblant à celle du criquet en vol. Cette découverte a été faite par Wilson en 1961. Depuis lors, des preuves de la présence de générateurs de motifs centraux chez les animaux vertébrés ont été apportées, à commencer par les travaux sur le chat dans les années 1960 d'Elzbieta Jankowska à Göteborg, qui a fourni la première preuve de l'existence d'un CPG de la moelle épinière. Cette section aborde le rôle du générateur de motifs centraux dans la locomotion de la lamproie et de l'homme.
Français La lamproie a été utilisée comme modèle pour les CPG des vertébrés car, bien que son système nerveux ait une organisation vertébrée, il partage de nombreuses caractéristiques positives avec les invertébrés. Lorsqu'elle est retirée de la lamproie, la moelle épinière intacte peut survivre pendant des jours in vitro . Elle a également très peu de neurones et peut être facilement stimulée pour produire un mouvement de nage fictif indiquant un générateur de motifs central. Dès 1983, Ayers, Carpenter, Currie et Kinch ont suggéré qu'il existait un CPG responsable de la plupart des mouvements ondulants chez la lamproie, y compris la nage en avant et en arrière, l'enfouissement dans la boue et le ramper sur une surface solide, qui, bien que sans surprise, ne corresponde pas à l'activité de l'animal intact, fournit néanmoins la sortie locomotrice de base. On a découvert que les différents mouvements étaient altérés par des neuromodulateurs, dont la sérotonine dans une étude de Harris-Warrick et Cohen en 1985 et la tachykinine dans une étude de Parker et al. en 1998. Le modèle de CPG de la lamproie pour la locomotion a été important pour l'étude des CPG. Bien que Sten Grillner affirme que le réseau locomoteur est caractérisé, une affirmation qui a apparemment été acceptée sans critique par le domaine du réseau locomoteur de la moelle épinière, il manque en fait de nombreux détails et Grillner ne peut pas fournir les preuves qu'il utilise pour étayer ses affirmations (Parker 2006). Cependant, ce modèle de circuit neuronal du CPG de la lamproie, comprenant trois classes (une excitatrice et deux inhibitrices) de neurones mais omettant les détails sous-cellulaires, fournit une compréhension au niveau du système de la locomotion générée par le CPG dont la vitesse et la direction (nager vers l'avant, vers l'arrière ou tourner) sont définies par des entrées externes non rythmiques (du tronc cérébral ) au circuit. Un schéma général du CPG de la lamproie est maintenant utilisé dans la création de CPG artificiels. Par exemple, Ijspeert et Kodjabachian ont utilisé le modèle d'Ekeberg pour la lamproie afin de créer des CPG artificiels et de simuler des mouvements de nage dans un substrat de type lamproie à l'aide de contrôleurs basés sur un codage SGOCE. Il s'agit essentiellement des premières étapes vers l'utilisation de CPG pour coder la locomotion chez les robots. Le modèle vertébré de CPG a également été développé avec le formalisme de Hodgkin-Huxley, ses variantes et les approches de systèmes de contrôle. Par exemple, Yakovenko et ses collègues ont développé un modèle mathématique simple qui décrit les principes de base proposés par TG Brown avec des unités d'intégration au seuil organisées avec des connexions mutuellement inhibitrices. Ce modèle est suffisant pour décrire des propriétés complexes du comportement, telles que les différents régimes de locomotion dominante en extenseur et en flexion observés lors de la stimulation électrique de la région locomotrice mésencéphalique (RLM), la locomotion fictive induite par la RLM.
Français Les connexions entre les CPG qui contrôlent chaque membre gèrent la coordination entre les membres et donc les démarches chez les animaux quadrupèdes et peut-être aussi bipèdes. La coordination gauche-droite est médiée par les commissures et l'avant-arrière ainsi que la coordination diagonale est médiée par les interneurones propiospinaux à longue projection. L'équilibre de l'alternance gauche-droite (classes de neurones V0d et V0v identifiées génétiquement médiées) aux interneurones commissuraux favorisant la synchronisation gauche (neurones V3 potentiellement médiés) détermine si la marche et le trot (allures alternées) ou le galop et le bond (allures synchrones) sont exprimés. Cet équilibre change avec l'augmentation de la vitesse, potentiellement en raison de la modulation par la commande supraspinale du MLR et médiée par la formation réticulaire, et provoque des transitions de démarche dépendantes de la vitesse caractéristiques des animaux quadrupèdes. La transition de la marche au trot se produit potentiellement en raison de la diminution plus forte des durées de la phase d'extension que de la phase de flexion avec l'augmentation de la vitesse locomotrice et pourrait être médiée par une inhibition diagonale descendante via les neurones propriospinaux longs V0d, ce qui conduit à un chevauchement progressivement croissant entre les membres diagonaux jusqu'à la synchronisation diagonale (trot). Les neurones commissuraux et propriospinaux longs sont une cible probable des entrées afférentes supraspinales et somatosensorielles pour ajuster la coordination entre les membres et la démarche à différentes conditions environnementales et comportementales.
Les générateurs de rythmes centraux contribuent également à la locomotion chez l'homme. En 1994, Calancie et al. ont décrit le « premier exemple bien défini d'un générateur de rythme central pour la marche chez l'adulte humain ». Le sujet était un homme de 37 ans qui avait subi une blessure à la moelle épinière cervicale 17 ans auparavant. Après une paralysie totale initiale sous le cou, le sujet a finalement retrouvé une certaine mobilité des bras et des doigts et une mobilité limitée des membres inférieurs. Il n'avait pas suffisamment récupéré pour supporter son propre poids. Après 17 ans, le sujet a constaté que lorsqu'il était allongé sur le dos et qu'il étendait ses hanches, ses membres inférieurs subissaient des mouvements de type marche tant qu'il restait allongé. « Les mouvements (i) impliquaient une flexion et une extension alternées de ses hanches, de ses genoux et de ses chevilles ; (ii) étaient fluides et rythmés ; (iii) étaient suffisamment puissants pour que le sujet se sente rapidement mal à l'aise en raison d'une « tension » musculaire excessive et d'une température corporelle élevée ; et (iv) ne pouvaient pas être arrêtés par un effort volontaire. » Après une étude approfondie du sujet, les expérimentateurs ont conclu que « ces données représentent la preuve la plus claire à ce jour qu'un tel réseau [CPG] existe chez l'homme. » Quatre ans plus tard, en 1998, Dimitrijevic et al. ont montré que les réseaux générateurs de motifs lombaires humains peuvent être activés par une commande vers des afférences sensorielles de grand diamètre des racines postérieures. Lorsqu'une stimulation électrique tonique est appliquée à ces fibres chez des individus atteints d'une lésion motrice complète de la moelle épinière (c'est-à-dire des individus chez qui la moelle épinière est fonctionnellement isolée du cerveau), des mouvements rythmiques de type locomoteur des membres inférieurs peuvent être provoqués. Ces mesures ont été effectuées en position couchée, minimisant ainsi la rétroaction périphérique. Des études ultérieures ont montré que ces centres locomoteurs lombaires peuvent former une grande variété de mouvements rythmiques en combinant et en distribuant des motifs stéréotypés aux nombreux muscles des membres inférieurs. Un traitement médicamenteux activant la CPG appelé Spinalon, actif de manière centrale après administration orale, a également montré qu'il réactivait partiellement les neurones locomoteurs spinaux chez les patients présentant une lésion médullaire complète ou motrice complète. En effet, une étude randomisée en double aveugle contrôlée par placebo menée auprès de quarante-cinq volontaires atteints de lésions chroniques de la moelle épinière A/B (entre 3 mois et 30 ans après le traumatisme) et couchés en décubitus dorsal pour des raisons de sécurité, a révélé que le Spinalon en dessous de la dose maximale tolérée (la dose maximale tolérée était de 500/125/50 mg/kg de L-DOPA/carbidopa/buspirone) était bien toléré. Des preuves préliminaires d'efficacité ont également été trouvées en utilisant des enregistrements vidéo et électromyographiques puisque des doses inférieures à la dose maximale tolérée pouvaient induire de manière aiguë des mouvements rythmiques des jambes de type locomoteur dans les groupes sous Spinalon, mais pas dans ceux sous placebo (fécule de maïs).
Contrôle de la locomotion
Si les durées des cycles de pas et les activations musculaires étaient fixes, il ne serait pas possible de modifier la vitesse du corps et de s'adapter à des terrains variés. Il a été suggéré que le CPG locomoteur des mammifères comprend un « minuteur » (peut-être sous la forme d'oscillateurs couplés) qui génère des cycles de pas de durées variables, et une « couche de formation de motifs », qui sélectionne et classe l'activation des pools moteurs . L'augmentation de la commande neurale de la région locomotrice du mésencéphale (MLR) au CPG spinal augmente la fréquence du cycle de pas (la cadence). Les durées des phases de balancement et d'appui co-varient dans une relation assez fixe, les phases d'appui changeant plus que les phases de balancement.
Les entrées sensorielles des membres peuvent tronquer ou prolonger les durées de phase individuelles dans un processus apparenté au contrôle d'état fini (dans lequel les règles « si-alors » déterminent quand les transitions d'état se produisent). Par exemple, si un membre qui se balance vers l'avant atteint la fin du balancement en moins de temps que la durée actuelle de la phase de flexion générée par le CPG, l'entrée sensorielle amènerait le minuteur CPG à terminer le balancement et à démarrer la phase d'appui. De plus, à mesure que la vitesse du corps augmente, la couche de formation de motifs augmenterait l'activation musculaire de manière non linéaire pour fournir des forces de charge et de poussée accrues. Il a été postulé que dans les mouvements bien prédits, les durées de phase générées par le CPG et les forces musculaires correspondent étroitement à celles requises par les événements biomécaniques en évolution, minimisant les corrections sensorielles requises. Le terme « réglage neuromécanique » a été inventé pour décrire ce processus.
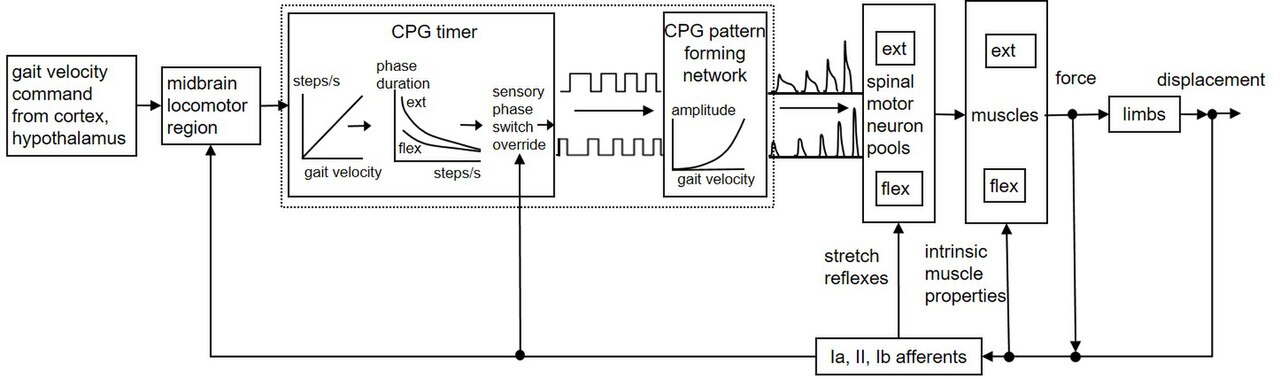
La figure 1 fournit un schéma simplifié qui résume ces mécanismes proposés. Une commande spécifiant la vitesse corporelle souhaitée descend des centres supérieurs vers le MLR, qui pilote le CPG locomoteur spinal. Le minuteur CPG produit la cadence et les durées de phase appropriées et la couche de formation de motifs module les sorties motoneuronales. Les muscles activés résistent à l'étirement grâce à leurs propres propriétés biomécaniques intrinsèques, fournissant une forme rapide de contrôle par rétroaction de la longueur et de la vitesse. Les réflexes médiés par l'organe tendineux de Golgi et d'autres afférents fournissent une compensation de charge supplémentaire, mais le rôle principal de l'entrée sensorielle peut être d'ajuster ou de remplacer le CPG lors des transitions position-balancement-position.
Comme décrit dans Neuromodulation, le CPG locomoteur humain est très adaptable et peut répondre aux entrées sensorielles. Il reçoit des informations du tronc cérébral ainsi que de l'environnement pour maintenir le réseau régulé. Des études plus récentes ont non seulement confirmé la présence du CPG pour la locomotion humaine, mais ont également confirmé sa robustesse et son adaptabilité. Par exemple, Choi et Bastian ont montré que les réseaux responsables de la marche humaine sont adaptables sur des échelles de temps courtes et longues. Ils ont montré une adaptation à différents modèles de marche et à différents contextes de marche. Ils ont également montré que différents modèles moteurs peuvent s'adapter indépendamment. Les adultes pourraient même marcher sur des tapis roulants en allant dans une direction différente pour chaque jambe. Cette étude a montré que des réseaux indépendants contrôlent la marche en avant et en arrière et que les réseaux contrôlant chaque jambe peuvent s'adapter indépendamment et être entraînés à marcher indépendamment. Ainsi, les humains possèdent également un générateur de modèles central pour la locomotion qui est capable non seulement de générer des modèles rythmiques mais aussi d'une adaptation et d'une utilité remarquables dans une grande variété de situations.
Respiration
Le modèle classique du CPG respiratoire est un modèle triphasé. Les phases du CPG respiratoire sont caractérisées par l'activité rythmique : (1) du nerf phrénique pendant l'inspiration ; (2) des branches du nerf laryngé récurrent qui innervent le muscle thyro-aryténoïdien pendant la dernière étape de l'expiration ; (3) des branches du nerf intercostal interne qui innervent le muscle triangularis sterni pendant la deuxième étape de l'expiration. La rythmicité de ces nerfs est classiquement considérée comme provenant d'un seul générateur de rythme. Dans ce modèle, la mise en phase est produite par une inhibition synaptique réciproque entre des groupes d'interneurones actifs séquentiellement.
Néanmoins, un modèle alternatif a été proposé renforcé par certaines données expérimentales. Selon ce modèle, le rythme respiratoire est généré par deux générateurs de rythme anatomiquement distincts couplés, l'un dans le complexe pré-Boetzinger et l'autre dans le noyau rétrotrapézoïdal / groupe respiratoire parafacial . Une étude plus approfondie a apporté des preuves à l'hypothèse selon laquelle l'un des réseaux est responsable du rythme d'inspiration et l'autre du rythme d'expiration. Par conséquent, l'inspiration et l'expiration sont des fonctions distinctes et l'une n'induit pas l'autre, comme on le croit communément, mais l'une des deux domine le comportement en générant un rythme plus rapide.
Déglutition
La déglutition implique la contraction coordonnée de plus de 25 paires de muscles de l'oropharynx, du larynx et de l'œsophage, qui sont actifs pendant une phase oropharyngée, suivie du péristaltisme œsophagien primaire. La déglutition dépend d'un centre de déglutition , un CPG situé dans la moelle allongée , qui implique plusieurs noyaux moteurs du tronc cérébral et deux groupes principaux d'interneurones : un groupe de déglutition dorsal (DSG) dans le noyau du tractus solitaire et un groupe de déglutition ventral (VSG) situé dans la moelle ventrolatérale au-dessus du noyau ambigu. Les neurones du DSG sont responsables de la génération du schéma de déglutition, tandis que ceux du VSG distribuent les commandes aux différents pools motoneuronaux. Comme dans les autres CPG, le fonctionnement du réseau central peut être modulé par des entrées périphériques et centrales, de sorte que le schéma de déglutition est adapté à la taille du bol alimentaire.
Au sein de ce réseau, les connexions inhibitrices centrales jouent un rôle majeur, produisant une inhibition rostrocaudale qui correspond à l'anatomie rostrocaudale du tractus de déglutition. Ainsi, lorsque les neurones contrôlant les parties proximales du tractus sont actifs, ceux qui commandent les parties plus distales sont inhibés. Outre le type de connexion entre les neurones, les propriétés intrinsèques des neurones, en particulier celles des neurones NTS, contribuent probablement également à la formation et au timing du schéma de déglutition.
Le CPG de déglutition est un CPG flexible. Cela signifie qu'au moins certains des neurones de déglutition peuvent être des neurones multifonctionnels et appartenir à des pools de neurones communs à plusieurs CPG. L'un de ces CPG est le CPG respiratoire, qui a été observé en interaction avec le CPG de déglutition.
Générateurs de rythme
Les générateurs de motifs centraux peuvent également jouer un rôle dans la génération de rythme pour d'autres fonctions chez les vertébrés. Par exemple, le système vibrisse du rat utilise un CPG non conventionnel pour les mouvements de fouet . « Comme d'autres CPG, le générateur de fouet peut fonctionner sans apport cortical ou rétroaction sensorielle. Cependant, contrairement à d'autres CPG, les motoneurones vibrisses participent activement à la rythmogenèse en convertissant les apports sérotoninergiques toniques en sortie motrice structurée responsable du mouvement des vibrisses. » La respiration est une autre fonction non locomotrice des générateurs de motifs centraux. Par exemple, les larves d'amphibiens accomplissent l'échange de gaz en grande partie grâce à la ventilation rythmique des branchies. Une étude a montré que la ventilation pulmonaire dans le tronc cérébral du têtard peut être pilotée par un mécanisme de type stimulateur cardiaque, tandis que le CPG respiratoire s'adapte chez la grenouille-taureau adulte à mesure qu'elle mûrit. Ainsi, les CPG remplissent une large gamme de fonctions chez l'animal vertébré et sont largement adaptables et variables avec l'âge, l'environnement et le comportement.
Mécanisme
La rythmicité des CPG peut également résulter de propriétés cellulaires dépendantes du temps, telles que l'adaptation, l'excitation retardée et le rebond post-inhibiteur (PIR). Le PIR est une propriété intrinsèque qui provoque une activité électrique rythmique en dépolarisant la membrane une fois que le stimulus hyperpolarisant a disparu. Il peut être produit par plusieurs mécanismes, notamment le courant cationique activé par hyperpolarisation (Ih), le courant calcique activé à basse tension ou la désactivation des courants entrants activés par dépolarisation . Une fois l'inhibition terminée, cette période de PIR peut être expliquée comme la période d'excitabilité neuronale accrue. C'est la propriété de nombreux neurones du SNC qui entraîne parfois des « explosions » de potentiel d'action immédiatement après une entrée synaptique inhibitrice. « Pour cette raison, il a été suggéré que le PIR peut contribuer au maintien de l'activité oscillatoire dans les réseaux neuronaux caractérisés par des connexions inhibitrices mutuelles, comme celles impliquées dans les comportements locomoteurs. De plus, le PIR est souvent inclus comme élément dans les modèles informatiques de réseaux neuronaux qui impliquent une inhibition mutuelle ». Par exemple, le « PIR dans les neurones récepteurs d'étirement des écrevisses est causé par la récupération de l'adaptation au cours de l'hyperpolarisation inhibitrice. Une caractéristique de ce système est que le PIR ne se produit que si l'hyperpolarisation est imposée sur un fond d'excitation, provoquée dans ce cas par l'étirement. Ils ont également découvert que le PIR peut être provoqué dans le récepteur d'étirement par des impulsions de courant hyperpolarisantes. Il s'agissait d'une découverte importante car elle montrait que le PIR est une propriété intrinsèque du neurone postsynaptique, liée au changement de potentiel de membrane associé à l'inhibition mais indépendante des récepteurs transmetteurs ou des propriétés présynaptiques. Cette dernière conclusion a résisté à l'épreuve du temps, faisant du PIR une propriété robuste des neurones du SNC dans une grande variété de contextes. » Cette propriété cellulaire est plus facilement observable dans le circuit neuronal de la lamproie. Le mouvement de nage est produit par une activité neuronale alternée entre le côté gauche et le côté droit du corps, ce qui provoque une flexion d'avant en arrière tout en créant des mouvements oscillatoires. Alors que la lamproie est penchée vers la gauche, il y a une inhibition réciproque du côté droit qui provoque une relaxation due à l'hyperpolarisation. Immédiatement après ce stimulus hyperpolarisant, les interneurones utilisent le rebond post-inhibiteur pour initier l'activité du côté droit. La dépolarisation de la membrane provoque sa contraction tandis que l'inhibition réciproque est maintenant appliquée au côté gauche.
Fonctions chez les invertébrés
Les CPG jouent un rôle tout aussi essentiel dans la coordination des comportements chez les invertébrés, et l'étude des CPG des invertébrés avec un nombre réduit de neurones a aidé à établir les principes généraux des CPG et de leur organisation dans le système nerveux. Un circuit modèle pour l'étude des CPG est le ganglion stomatogastrique des crabes et des homards, un circuit d'environ 30 neurones contenant deux CPG qui génèrent une sortie motrice rythmique pour la mastication et la digestion des aliments. La dissection de ces circuits a révélé les mécanismes neuronaux des CPG. Par exemple, le CPG pylorique - qui contrôle la contraction et la dilatation du pylore - contient un ensemble de neurones oscillatoires conditionnels et un neurone stimulateur cardiaque qui s'active de manière rythmique lorsqu'il est disséqué hors du circuit. Les comportements rythmiques coordonnés comme la marche, le vol et le toilettage sont également contrôlés par les CPG chez certains invertébrés. Des recherches continues sur la façon dont les CPG contrôlent ces comportements ont révélé une architecture de CPG imbriquée pour contrôler les comportements rythmiques sur différentes échelles de temps. D'autres exemples de CPG chez les animaux invertébrés comprennent un CPG modulant le retrait réflexe, la nage de fuite et la reptation chez le mollusque Tritonia, et pour contrôler le rythme cardiaque des sangsues. Les générateurs de motifs centraux jouent un rôle important chez tous les animaux et présentent une variabilité et une adaptabilité étonnantes dans presque tous les cas.
Interprétations alternatives
Une théorie qui réconcilie le rôle du retour sensoriel pendant la locomotion rythmique consiste à redéfinir les CPG comme des « estimateurs d'état » par opposition aux générateurs de rythme. Dans cette perspective, les CPG sont un processeur spinal intrinsèque qui corrige le retour sensoriel imparfait et adapte l'entrée centrale à cette entrée périphérique optimisée. Les modèles utilisant ce cadre sont capables d'accomplir un comportement rythmique ainsi qu'une locomotion fictive sans incorporer de générateurs de rythme indépendants.