Le facteur d'initiation eucaryote 2 ( eIF2 ) est un facteur d'initiation eucaryote . Il est nécessaire à la plupart des formes d'initiation de la traduction eucaryote . L'eIF2 assure la liaison de l'ARNt i Met au ribosome d'une manière dépendante du GTP . L'eIF2 est un hétérotrimère constitué d'une sous-unité alpha (également appelée sous-unité 1, EIF2S1), d'une sous- unité bêta (sous-unité 2, EIF2S2) et d'une sous-unité gamma (sous-unité 3, EIF2S3).
Une fois la phase d'initiation terminée, eIF2 est libéré du ribosome lié au GDP sous forme de complexe binaire inactif. Pour participer à un autre cycle d'initiation de la traduction, ce GDP doit être échangé contre du GTP.
Fonction
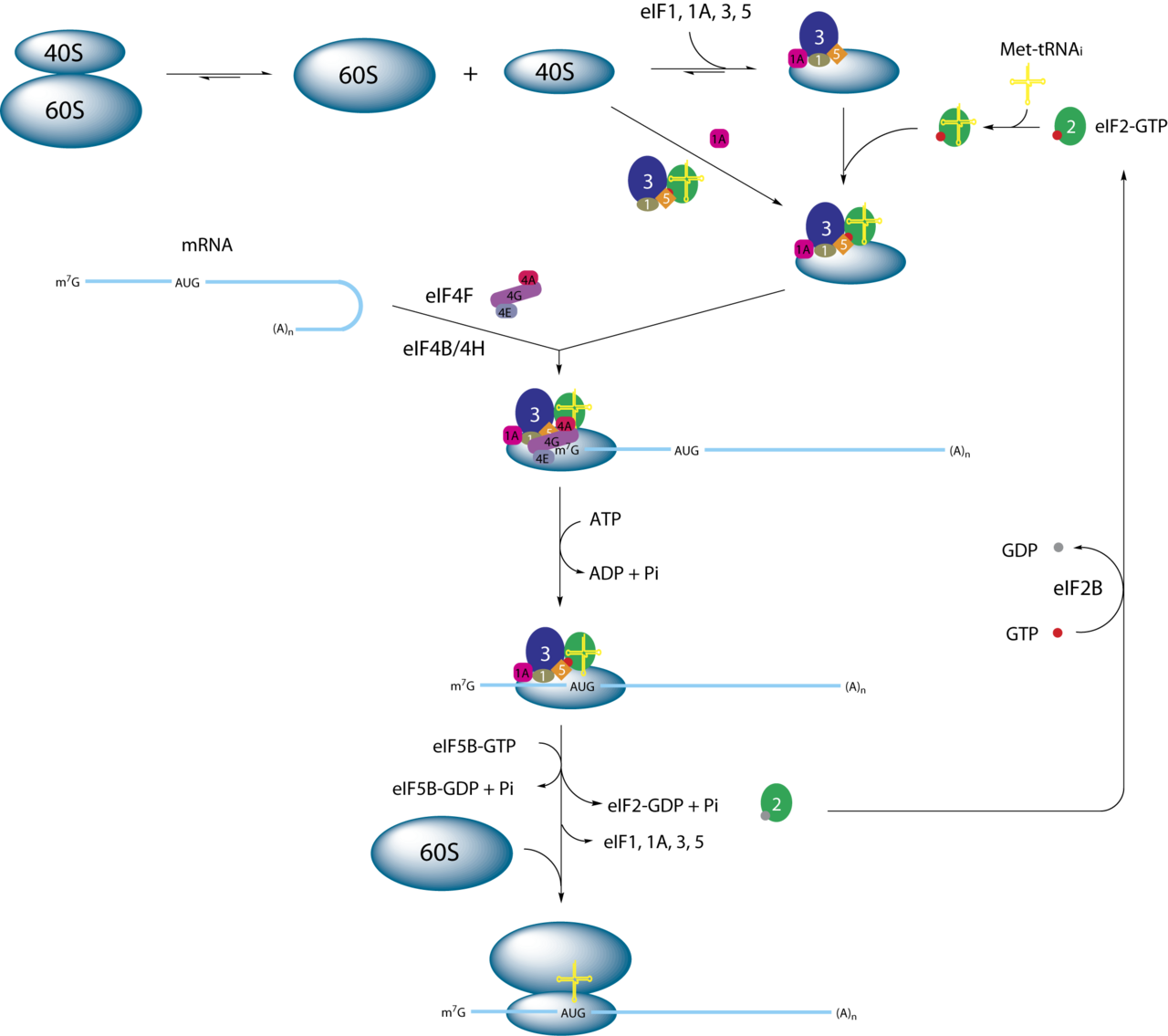
eIF2 est un facteur essentiel pour la synthèse des protéines qui forme un complexe ternaire (TC) avec le GTP et l'initiateur Met - ARNt i Met . Après sa formation, le TC se lie à la sous-unité ribosomique 40S pour former le complexe de préinitiation 43S (43S PIC). On pense que l'assemblage du 43S PIC est stimulé par les facteurs d'initiation eIF1 , eIF1A et le complexe eIF3 selon des expériences in vitro . Le 43S PIC se lie ensuite à l'ARNm qui a été précédemment déroulé par le complexe eIF4F . Les protéines 43S PIC et eIF4F forment un nouveau complexe 48S sur l'ARNm, qui commence à rechercher le long de l'ARNm le codon de départ (AUG). Lors de l'appariement des bases du codon AUG avec le Met-ARNt, eIF5 (qui est une protéine activatrice de GTPase , ou GAP) est recrutée dans le complexe et induit eIF2 à hydrolyser son GTP. Cela provoque la libération de eIF2-GDP de ce complexe 48S et la traduction commence après le recrutement de la sous-unité ribosomique 60S et la formation du complexe d'initiation 80S . Enfin, avec l'aide du facteur d'échange de nucléotides guanine (GEF) eIF2B , le GDP dans eIF2 est échangé contre un GTP et le complexe ternaire se reforme pour un nouveau cycle d'initiation de la traduction.
Structure
eIF2 est un hétérotrimère d'une masse molaire totale de 126 kDa composé de trois sous-unités : α (sous-unité 1), β (sous-unité 2) et γ (sous-unité 3). Les séquences des trois sous-unités sont hautement conservées (les identités d'acides aminés par paire pour chaque sous-unité varient de 47 à 72 % en comparant les protéines d' Homo sapiens et de Saccharomyces cerevisiae ).
La sous-unité α contient la principale cible de phosphorylation , une sérine en position 51. Elle contient également un domaine de motif S1, qui est un site potentiel de liaison de l'ARN. Par conséquent, la sous-unité α peut être considérée comme la sous-unité régulatrice du trimère.
La sous-unité β contient plusieurs sites de phosphorylation (résidus 2, 13, 67, 218). Il est important de prendre en compte qu'il existe également trois groupes de lysine dans le domaine N-terminal (NTD), qui sont importants pour l'interaction avec eIF2B. De plus, la séquence de la protéine comprend un motif de doigt de zinc qui s'est avéré jouer un rôle dans la formation du complexe ternaire et du complexe de préinitiation 43S. Il existe également deux séquences de liaison aux nucléotides de guanine qui ne se sont pas révélées impliquées dans la régulation de l'activité d'eIF2. On pense également que la sous-unité β interagit à la fois avec l'ARNt et l'ARNm.
La sous-unité γ comprend trois sites de liaison aux nucléotides guanine et est connue pour être le principal site d'amarrage du GTP/GDP. Elle contient également une cavité de liaison à l'ARNt qui a été mise en évidence par cristallographie aux rayons X. Un motif de jointure en zinc est capable de lier un cation Zn 2+ . Il est lié à certains facteurs d'élongation comme EF-Tu .
Règlement
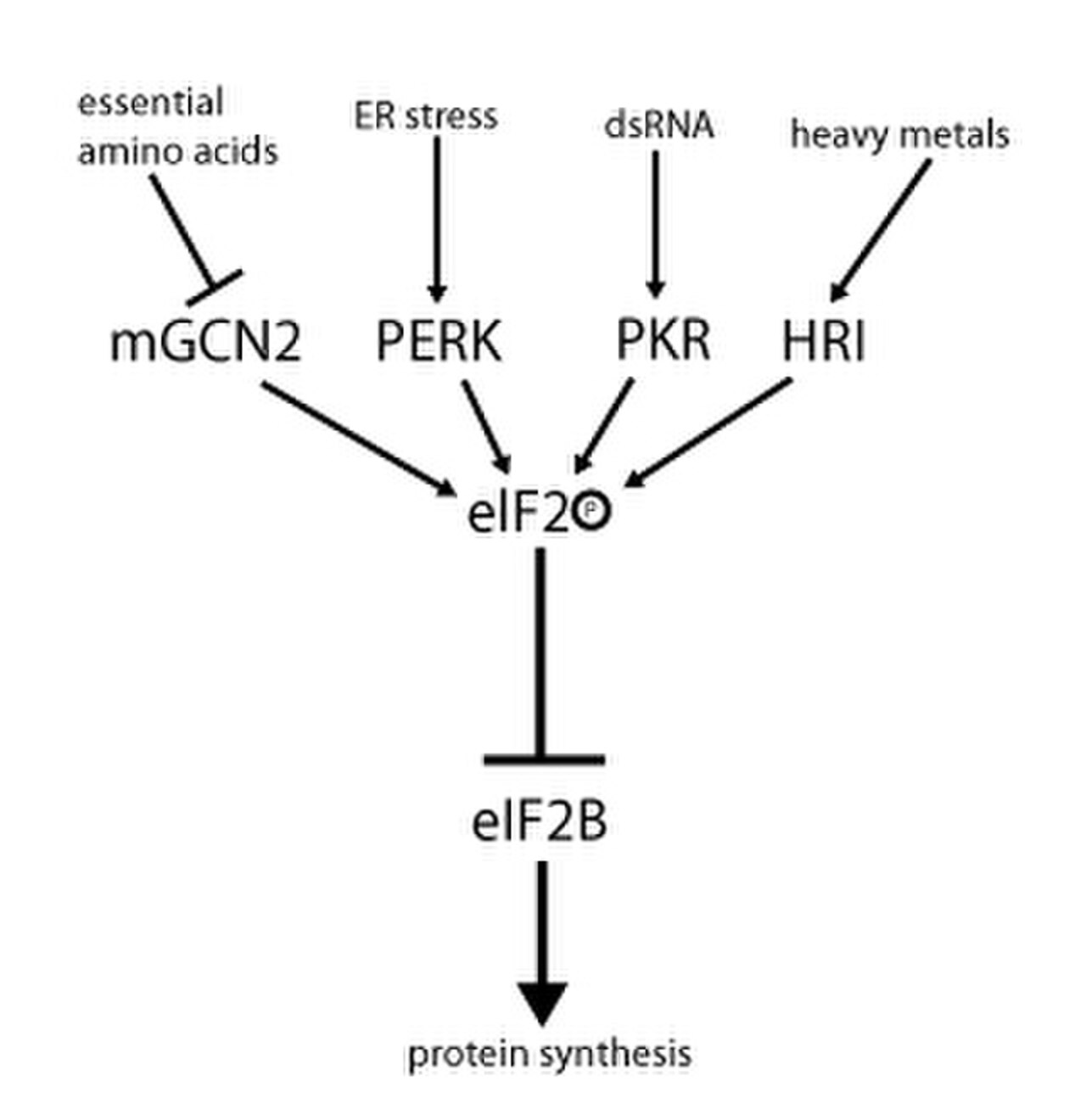
L'activité d'eIF2 est régulée par un mécanisme impliquant à la fois l'échange de nucléotides guanine et la phosphorylation. La phosphorylation a lieu au niveau de la sous-unité α, qui est une cible pour un certain nombre de sérine kinases qui phosphorylent la sérine 51. Ces kinases agissent en raison d'un stress tel que la privation d'acides aminés ( GCN2 ), le stress du RE ( PERK ), la présence d'un déficit en hème dsRNA ( PKR ) ( HRI ) ou l'interféron . Une fois phosphorylé, eIF2 montre une affinité accrue pour eIF2B, son GEF. Cependant, eIF2B est capable d'échanger GDP contre GTP uniquement si eIF2 est dans son état non phosphorylé. Cependant, eIF2 phosphorylé, en raison de sa liaison plus forte, agit comme un inhibiteur de son propre GEF (eIF2B). Étant donné que la concentration cellulaire d'eIF2B est bien inférieure à celle d'eIF2, même une petite quantité d'eIF2 phosphorylé peut complètement abolir l'activité d'eIF2B par séquestration. Sans le GEF, eIF2 ne peut plus revenir à son état actif (lié au GTP). En conséquence, la traduction s'arrête puisque l'initiation n'est plus possible sans aucun complexe ternaire disponible. De plus, une faible concentration de complexe ternaire permet l'expression de GCN4 (condition de famine), ce qui, à son tour, entraîne une activation accrue des gènes de synthèse des acides aminés
Maladie
Étant donné que l'eIF2 est essentielle à la plupart des formes d'initiation de la traduction et donc à la synthèse des protéines, les défauts de l'eIF2 sont souvent mortels. La protéine est hautement conservée parmi les espèces éloignées de l'évolution, ce qui indique un impact important des mutations sur la viabilité cellulaire. Par conséquent, aucune maladie directement liée aux mutations de l'eIF2 n'a pu être observée. Cependant, de nombreuses maladies sont causées par une régulation négative de l'eIF2 par ses kinases en amont. Par exemple, des concentrations accrues de PKR active et d'eIF2 inactive (phosphorylée) ont été trouvées chez des patients atteints de maladies neurodégénératives telles que la maladie d'Alzheimer , la maladie de Parkinson et la maladie de Huntington . Il existe également un exemple avéré d'une maladie liée au GEF eIF2B. Les mutations dans les cinq sous-unités de l'eIF2B sont associées à la maladie de la substance blanche disparaissante (VWM), une leucodystrophie génétique qui provoque la dégénérescence et la disparition de la substance blanche du cerveau. On ne comprend toujours pas complètement pourquoi seules les cellules cérébrales semblent être affectées par ces défauts. Des niveaux potentiellement réduits de protéines régulatrices instables pourraient jouer un rôle dans le développement des maladies mentionnées.