
La ligne latérale , également appelée organe de la ligne latérale ( OLL ), est un système d' organes sensoriels présent chez les poissons , utilisé pour détecter les mouvements, les vibrations et les gradients de pression dans l'eau environnante. La capacité sensorielle est obtenue via des cellules épithéliales modifiées , appelées cellules ciliées , qui répondent au déplacement causé par le mouvement et transforment ces signaux en impulsions électriques via des synapses excitatrices . Les lignes latérales jouent un rôle important dans le comportement de regroupement , la prédation et l'orientation.
Au début de l' évolution des poissons , certains organes sensoriels de la ligne latérale ont été modifiés pour fonctionner comme des électrorécepteurs appelés ampoules de Lorenzini . Le système de la ligne latérale est ancien et basal par rapport au clade des vertébrés, car on le retrouve chez les poissons qui ont divergé il y a plus de 400 millions d'années.
Fonction
Le système de ligne latérale permet de détecter les mouvements, les vibrations et les gradients de pression dans l'eau entourant un animal. Il joue un rôle essentiel dans l'orientation, la prédation et le regroupement des poissons en offrant une perception spatiale et la capacité de naviguer dans l'environnement. L'analyse a montré que le système de ligne latérale devrait être un système de détection passive efficace capable de distinguer les obstacles immergés par leur forme. La ligne latérale permet aux poissons de naviguer et de chasser dans des eaux avec une mauvaise visibilité.
Le système de ligne latérale permet aux poissons prédateurs de détecter les vibrations émises par leur proie et de s'orienter vers la source pour commencer l'action prédatrice. Les poissons prédateurs aveuglés restent capables de chasser, mais pas lorsque la fonction de la ligne latérale est inhibée par les ions cobalt .
La ligne latérale joue un rôle dans la formation des bancs de poissons. Les Pollachius virens aveuglés ont pu s'intégrer dans un banc, alors que les poissons dont les lignes latérales étaient coupées ne le pouvaient pas. Il se peut qu'elle ait évolué davantage pour permettre aux poissons de se nourrir dans des grottes sombres. Chez le poisson aveugle des grottes mexicain, Astyanax mexicanus , les neuromastes dans et autour de l' orbite de l'œil sont plus gros et environ deux fois plus sensibles que ceux des poissons vivant à la surface.
L'une des fonctions de la formation de bancs peut être de brouiller la ligne latérale des poissons prédateurs . Un seul poisson-proie crée un modèle simple de vitesse des particules , alors que les gradients de pression de nombreux poissons-proies nageant de près (en banc) se chevauchent, créant un modèle complexe. Il est donc difficile pour les poissons prédateurs d'identifier une proie individuelle grâce à la perception de la ligne latérale.
Anatomie
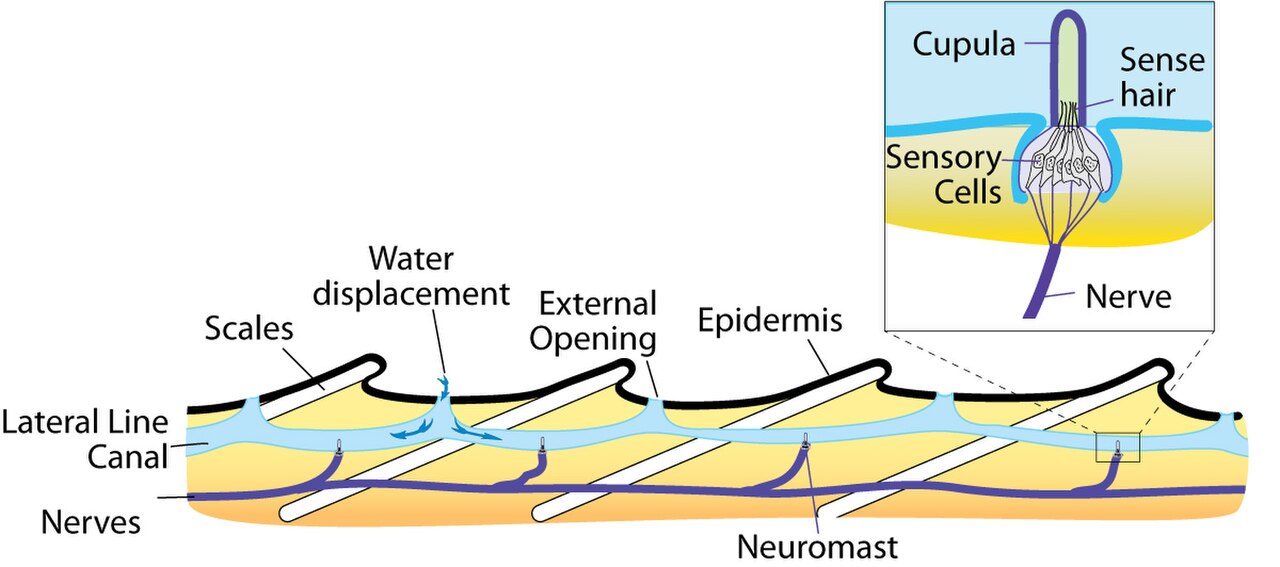

Les lignes latérales sont généralement visibles comme de faibles lignes de pores qui courent le long de chaque côté du corps d'un poisson. Les unités fonctionnelles de la ligne latérale sont les neuromastes, des organes mécano-récepteurs discrets qui détectent le mouvement dans l'eau. Il existe deux variétés principales : les neuromastes canalaires et les neuromastes superficiels. Les neuromastes superficiels se trouvent à la surface du corps, tandis que les neuromastes canalaires se trouvent le long des lignes latérales dans des canaux sous-cutanés remplis de liquide. Chaque neuromaste est constitué de cellules ciliées réceptrices dont les extrémités sont recouvertes d'une cupule gélatineuse flexible. Les cellules ciliées possèdent généralement à la fois des connexions afférentes glutamatergiques et des connexions efférentes cholinergiques . Les cellules ciliées réceptrices sont des cellules épithéliales modifiées ; elles possèdent généralement des faisceaux de 40 à 50 « poils » de microvillosités qui fonctionnent comme des mécanorécepteurs. Dans chaque faisceau, les poils sont organisés en un « escalier » grossier du plus court au plus long.
Transduction du signal
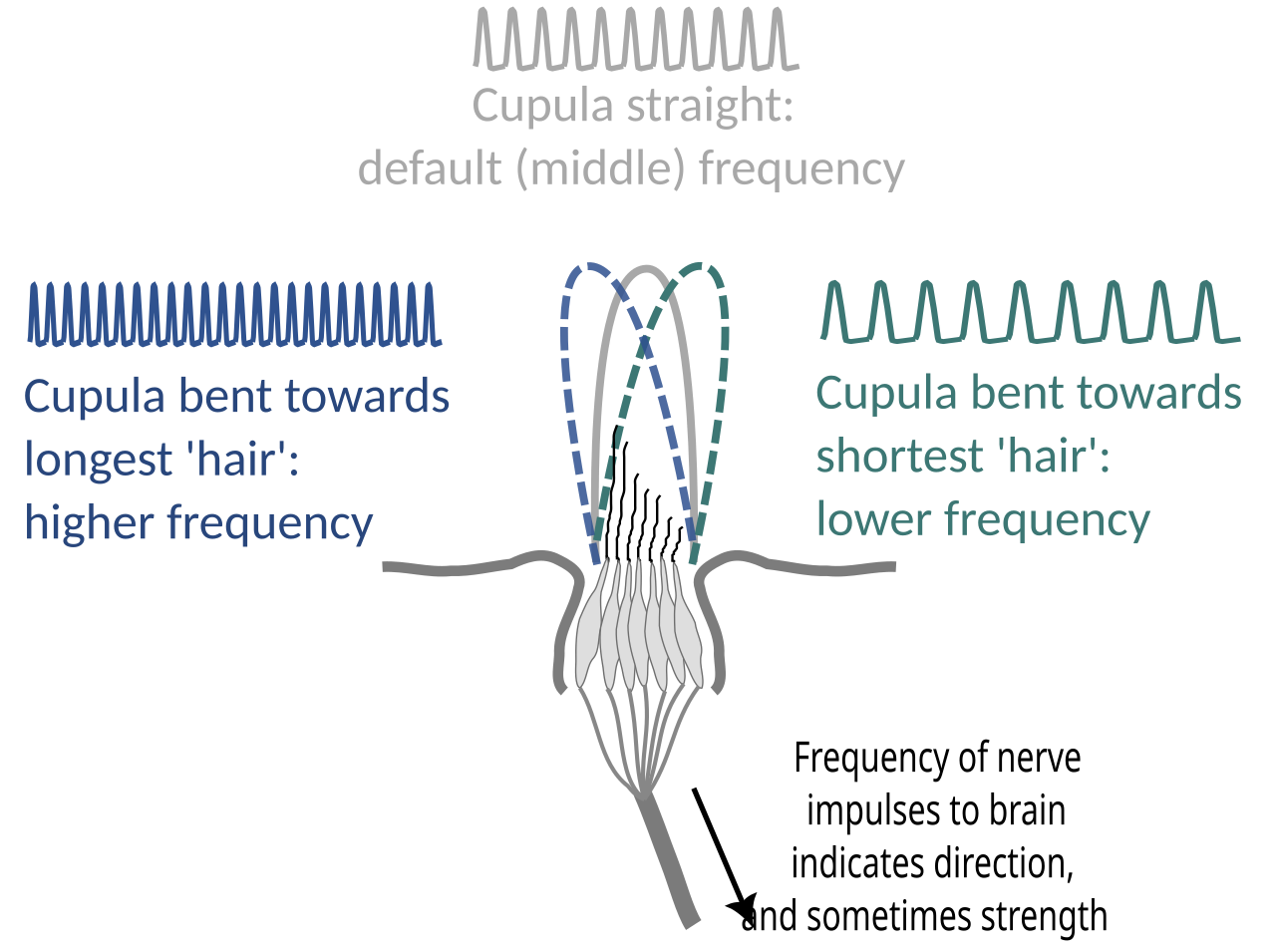
Les cellules ciliées sont stimulées par la déviation de leurs faisceaux de poils en direction des « poils » ou stéréocils les plus hauts . La déviation permet aux cations de pénétrer dans un canal mécaniquement fermé , provoquant une dépolarisation ou une hyperpolarisation de la cellule ciliée. La dépolarisation ouvre les canaux calciques Ca v 1.3 dans la membrane basolatérale .
Les cellules ciliées utilisent un système de transduction avec codage de vitesse pour transmettre la directionnalité d'un stimulus. Les cellules ciliées produisent une vitesse de décharge constante et tonique. Lorsque le mouvement mécanique est transmis par l'eau au neuromaste, la cupule se plie et se déplace en fonction de la force du stimulus. Cela entraîne un changement dans la perméabilité ionique de la cellule. La déviation vers le cheveu le plus long entraîne une dépolarisation de la cellule ciliée, une augmentation de la libération de neurotransmetteurs au niveau de la synapse afférente excitatrice et une augmentation du taux de transduction du signal . La déviation vers le cheveu le plus court a l'effet inverse, hyperpolarisant la cellule ciliée et produisant une diminution du taux de libération de neurotransmetteurs. Ces impulsions électriques sont ensuite transmises le long des neurones latéraux afférents au cerveau.
Bien que les deux variétés de neuromastes utilisent cette méthode de transduction, leur organisation spécialisée leur confère des capacités mécano-réceptrices différentes. Les organes superficiels sont exposés plus directement à l'environnement extérieur. L'organisation des faisceaux au sein de leurs organes est apparemment aléatoire, incorporant diverses formes et tailles de microvillosités au sein des faisceaux. Cela suggère une détection grossière mais de grande portée. En revanche, la structure des organes canalaires permet aux neuromastes canalaires une mécano-réception plus sophistiquée, comme celle des différences de pression. Lorsque le courant traverse les pores, une différence de pression est créée, induisant un écoulement dans le liquide canalaire. Cela déplace les cupules des neuromastes dans le canal, ce qui entraîne une déviation des poils dans la direction de l'écoulement.
Électrophysiologie
Les cellules ciliées mécano-réceptrices de la structure de la ligne latérale sont intégrées dans des circuits plus complexes par leurs connexions afférentes et efférentes. Les synapses qui participent directement à la transduction des informations mécaniques sont des connexions afférentes excitatrices qui utilisent le glutamate . Les espèces varient dans leurs neuromastes et leurs connexions afférentes, ce qui leur confère des propriétés mécano-réceptrices différentes. Par exemple, les neuromastes superficiels du poisson-aspirant , Porichthys notatus , sont sensibles à des fréquences de stimulation spécifiques. Une variété est adaptée pour recueillir des informations sur l'accélération, à des fréquences de stimulation comprises entre 30 et 200 Hz. L'autre type obtient des informations sur la vitesse et est plus réceptif à une stimulation inférieure à 30 Hz.
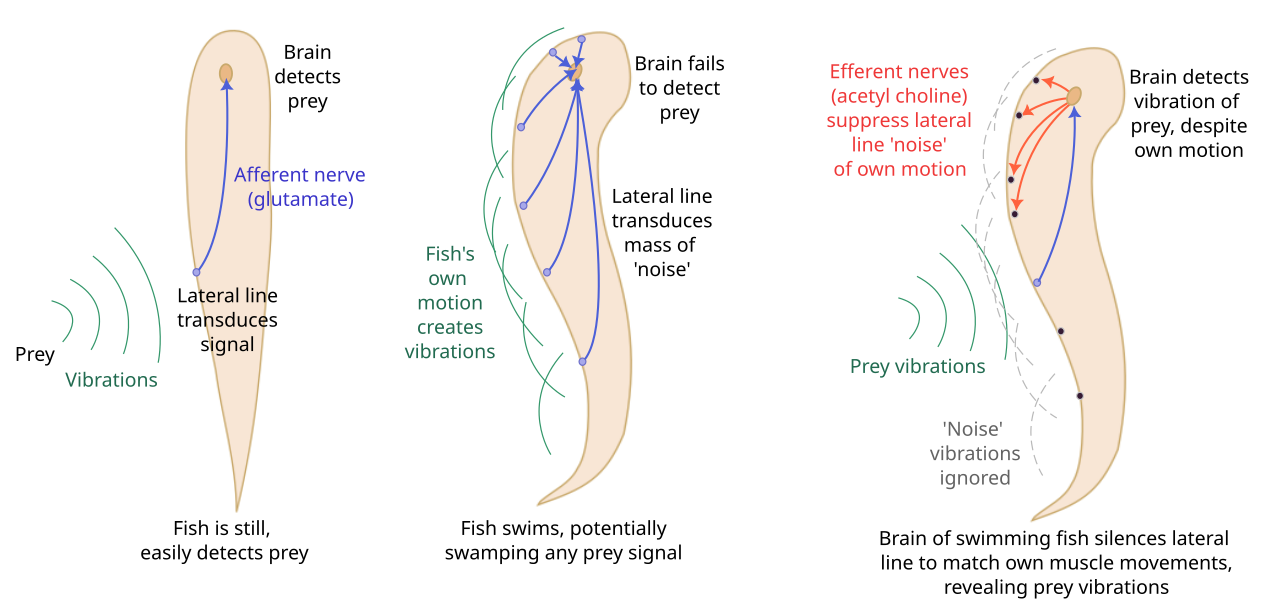
Les synapses efférentes des cellules ciliées sont inhibitrices et utilisent l'acétylcholine comme transmetteur. Elles sont des participants essentiels d'un système de décharge corollaire conçu pour limiter les interférences auto-générées. Lorsqu'un poisson se déplace, il crée des perturbations dans l'eau qui pourraient être détectées par le système de lignes latérales, interférant potentiellement avec la détection d'autres signaux biologiquement pertinents. Pour éviter cela, un signal efférent est envoyé à la cellule ciliée lors de l'action motrice, ce qui entraîne une inhibition qui contrecarre l'excitation résultant de la réception de la stimulation auto-générée. Cela permet au poisson de détecter les stimuli externes sans interférence de ses propres mouvements.
Les signaux des cellules ciliées sont transmis le long des neurones latéraux jusqu'au cerveau. La zone où ces signaux se terminent le plus souvent est le noyau octavolatéral médial (MON), qui traite et intègre probablement les informations mécano-réceptives. Le MON profond contient des couches distinctes de cellules de crête basilaires et non basilaires, suggérant des voies informatiques analogues au lobe latéral électrosensoriel des poissons électriques . Le MON est probablement impliqué dans l'intégration de circuits parallèles excitateurs et inhibiteurs pour interpréter les informations mécano-réceptives.
Évolution

L'utilisation de poils mécanosensibles est homologue au fonctionnement des cellules ciliées dans les systèmes auditif et vestibulaire , indiquant un lien étroit entre ces systèmes. En raison de nombreuses fonctions qui se chevauchent et de leur grande similitude en termes d'ultrastructure et de développement, le système de la ligne latérale et l' oreille interne des poissons sont souvent regroupés sous le nom de système octavolatéral (OLS). Ici, le système de la ligne latérale détecte les vitesses et les accélérations des particules avec des fréquences inférieures à 100 Hz. Ces basses fréquences créent de grandes longueurs d'onde, qui induisent de fortes accélérations des particules dans le champ proche des poissons nageurs qui ne rayonnent pas dans le champ lointain sous forme d'ondes acoustiques en raison d'un court-circuit acoustique . Le système auditif détecte les fluctuations de pression avec des fréquences supérieures à 100 Hz qui se propagent dans le champ lointain sous forme d'ondes.
Le système de la ligne latérale est ancien et basal par rapport au clade des vertébrés ; on le retrouve dans des groupes de poissons qui ont divergé il y a plus de 400 millions d'années, notamment les lamproies , les poissons cartilagineux et les poissons osseux . La plupart des larves d'amphibiens et certains amphibiens adultes entièrement aquatiques possèdent des systèmes mécanosensibles comparables à la ligne latérale. Les tétrapodes terrestres ont secondairement perdu leurs organes de ligne latérale, qui sont inefficaces lorsqu'ils ne sont pas immergés.
Les organes électrorécepteurs , appelés ampoules de Lorenzini , apparaissant comme des creux dans la peau des requins et de certains autres poissons, ont évolué à partir de l'organe de la ligne latérale. L'électroréception passive utilisant des ampoules est un trait ancestral chez les vertébrés, ce qui signifie qu'elle était présente chez leur dernier ancêtre commun .