Un motoneurone (ou motoneurone ou neurone efférent ) est un neurone dont le corps cellulaire est situé dans le cortex moteur , le tronc cérébral ou la moelle épinière , et dont l'axone (fibre) se projette vers la moelle épinière ou à l'extérieur de la moelle épinière pour contrôler directement ou indirectement les organes effecteurs, principalement les muscles et les glandes . Il existe deux types de motoneurones : les motoneurones supérieurs et les motoneurones inférieurs . Les axones des motoneurones supérieurs font synapse sur les interneurones de la moelle épinière et parfois directement sur les motoneurones inférieurs. Les axones des motoneurones inférieurs sont des fibres nerveuses efférentes qui transportent les signaux de la moelle épinière vers les effecteurs. Les types de motoneurones inférieurs sont les motoneurones alpha , les motoneurones bêta et les motoneurones gamma .
Un seul motoneurone peut innerver de nombreuses fibres musculaires et une fibre musculaire peut subir de nombreux potentiels d'action au cours d'une seule contraction musculaire . L'innervation a lieu au niveau d'une jonction neuromusculaire et les contractions peuvent se superposer à la suite d' une sommation ou d'une contraction tétanique . Les contractions individuelles peuvent devenir indiscernables et la tension augmente progressivement pour finalement atteindre un plateau.
Bien que le terme « motoneurone » suggère qu’il existe un seul type de neurone qui contrôle le mouvement, ce n’est pas le cas. En effet, les motoneurones supérieurs et inférieurs, qui diffèrent grandement par leur origine, la localisation des synapses, les voies, les neurotransmetteurs et les caractéristiques des lésions, sont inclus dans la même classification que les « motoneurones ». Essentiellement, les motoneurones, également appelés motoneurones, sont constitués d’une variété de circuits complexes et finement réglés présents dans tout le corps qui innervent les muscles et les glandes effecteurs pour permettre les mouvements volontaires et involontaires. Deux motoneurones se réunissent pour former un circuit à deux neurones. Alors que les motoneurones inférieurs commencent dans la moelle épinière et vont innerver les muscles et les glandes dans tout le corps, les motoneurones supérieurs naissent dans le cortex cérébral et se dirigent vers le tronc cérébral ou la moelle épinière. Il est essentiel de comprendre les distinctions entre les motoneurones supérieurs et inférieurs ainsi que les voies qu’ils suivent afin de détecter efficacement ces lésions neuronales et de localiser les lésions.
Développement
Les motoneurones commencent à se développer tôt dans le développement embryonnaire et la fonction motrice continue de se développer jusqu'à l'enfance. Dans le tube neural, les cellules sont spécifiées soit sur l'axe rostral-caudal, soit sur l'axe ventral-dorsal. Les axones des motoneurones commencent à apparaître au cours de la quatrième semaine de développement à partir de la région ventrale de l'axe ventral-dorsal (la plaque basale ). Cet homéodomaine est connu sous le nom de domaine progéniteur du motoneurone (pMN). Les facteurs de transcription ici incluent Pax6 , OLIG2 , Nkx-6.1 et Nkx-6.2 , qui sont régulés par Sonic Hedgehog (Shh). Le gène OLIG2 est le plus important en raison de son rôle dans la promotion de l'expression de Ngn2 , un gène qui provoque la sortie du cycle cellulaire ainsi que la promotion d'autres facteurs de transcription associés au développement du motoneurone.
Une spécification plus poussée des motoneurones se produit lorsque l'acide rétinoïque , le facteur de croissance des fibroblastes , les Wnts et le TGFb sont intégrés dans les différents facteurs de transcription Hox . Il existe 13 facteurs de transcription Hox et, avec les signaux, ils déterminent si un motoneurone aura un caractère plus rostral ou caudal. Dans la colonne vertébrale, les Hox 4 à 11 classent les motoneurones dans l'une des cinq colonnes motrices.
Anatomie et physiologie

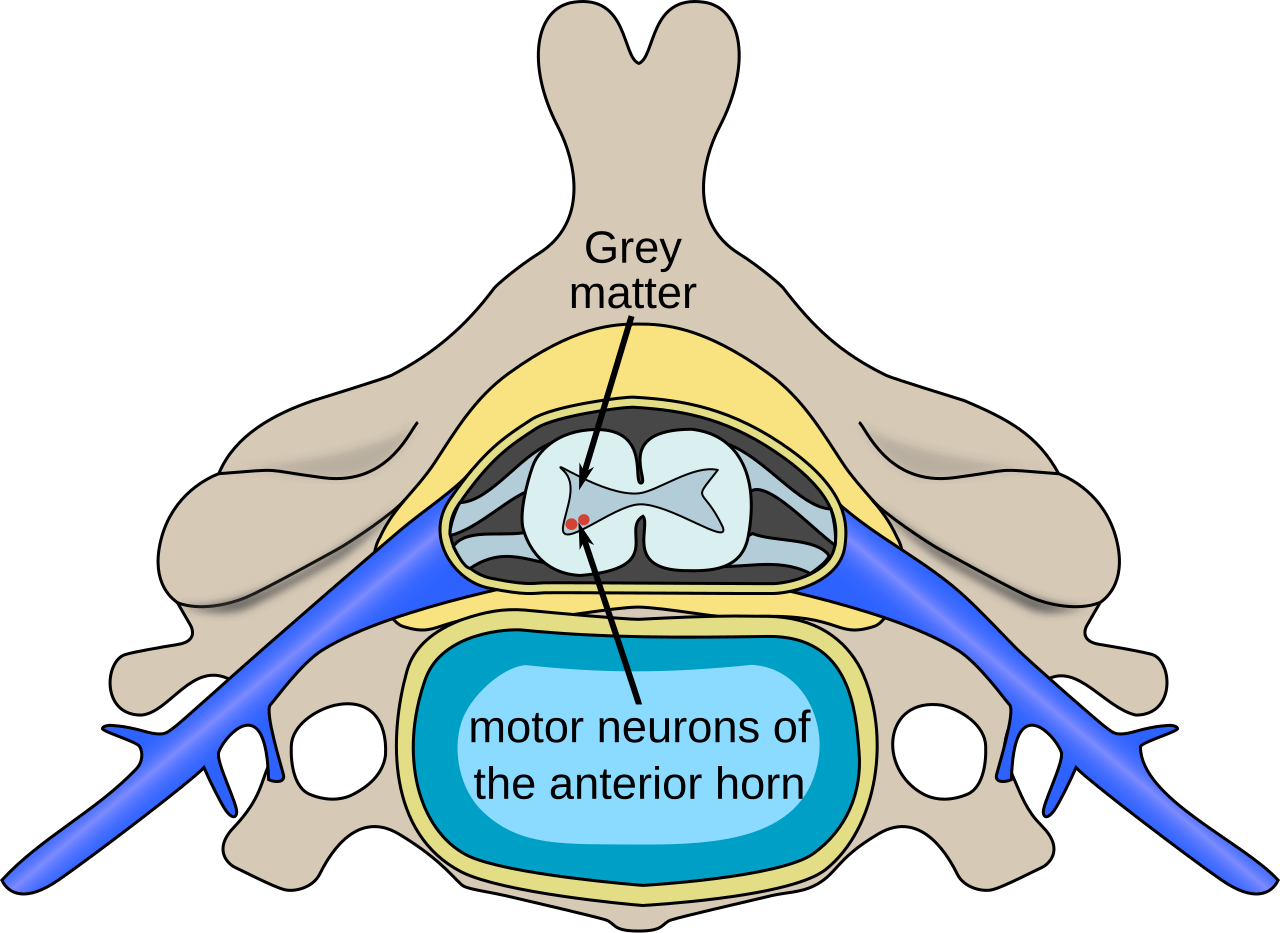
Motoneurones supérieurs
Les motoneurones supérieurs naissent dans le cortex moteur situé dans le gyrus précentral . Les cellules qui composent le cortex moteur primaire sont les cellules de Betz , qui sont des cellules pyramidales géantes . Les axones de ces cellules descendent du cortex pour former le tractus corticospinal . Les motoneurones corticospinaux se projettent du cortex primaire directement sur les motoneurones de la corne ventrale de la moelle épinière. Leurs axones font synapse sur les motoneurones spinaux de plusieurs muscles ainsi que sur les interneurones spinaux . Ils sont uniques aux primates et il a été suggéré que leur fonction est le contrôle adaptatif des mains, y compris le contrôle relativement indépendant des doigts individuels. Les motoneurones corticospinaux n'ont jusqu'à présent été trouvés que dans le cortex moteur primaire et non dans les zones motrices secondaires.
Voies nerveuses
Les voies nerveuses sont des faisceaux d'axones sous forme de matière blanche , qui transportent les potentiels d'action vers leurs effecteurs. Dans la moelle épinière, ces voies descendantes transportent des impulsions provenant de différentes régions. Ces voies servent également de lieu d'origine aux motoneurones inférieurs. On trouve sept voies motrices descendantes principales dans la moelle épinière :
- Voie corticospinale latérale
- Voie rubrospinale
- Voie réticulospinale latérale
- Voie vestibulo-spinale
- Voie réticulospinale médiale
- Voie tectospinale
- Voie corticospinale antérieure
Motoneurones inférieurs
Les motoneurones inférieurs sont ceux qui naissent dans la moelle épinière et innervent directement ou indirectement des cibles effectrices. La cible de ces neurones varie, mais dans le système nerveux somatique, la cible sera une sorte de fibre musculaire. Il existe trois catégories principales de motoneurones inférieurs, qui peuvent être subdivisées en sous-catégories.
Selon leurs cibles, les motoneurones sont classés en trois grandes catégories :
- Motoneurones somatiques
- Neurones moteurs viscéraux spéciaux
- Neurones moteurs viscéraux généraux
Motoneurones somatiques
Les neurones moteurs somatiques naissent dans le système nerveux central et projettent leurs axones vers les muscles squelettiques (tels que les muscles des membres, les muscles abdominaux et les muscles intercostaux ), qui sont impliqués dans la locomotion . Les trois types de ces neurones sont les neurones efférents alpha , les neurones efférents bêta et les neurones efférents gamma . Ils sont appelés efférents pour indiquer le flux d'informations du système nerveux central (SNC) vers la périphérie .
- Les neurones moteurs alpha innervent les fibres musculaires extrafusales , qui sont le principal composant générateur de force d'un muscle. Leurs corps cellulaires se trouvent dans la corne ventrale de la moelle épinière et sont parfois appelés cellules de la corne ventrale . Un seul neurone moteur peut faire synapse avec 150 fibres musculaires en moyenne. Le neurone moteur et toutes les fibres musculaires auxquelles il se connecte constituent une unité motrice . Les unités motrices sont divisées en 3 catégories :
- Les unités motrices lentes (S) stimulent les petites fibres musculaires, qui se contractent très lentement et fournissent de petites quantités d'énergie mais sont très résistantes à la fatigue, elles sont donc utilisées pour soutenir la contraction musculaire, comme le maintien du corps en position verticale. Elles obtiennent leur énergie par des moyens oxydatifs et nécessitent donc de l'oxygène. On les appelle également fibres rouges.
- Les unités motrices à fatigue rapide (FF) stimulent des groupes musculaires plus importants, qui appliquent de grandes quantités de force mais se fatiguent très rapidement. Elles sont utilisées pour des tâches qui nécessitent de grandes poussées d'énergie brèves, comme sauter ou courir. Elles obtiennent leur énergie par des moyens glycolytiques et ne nécessitent donc pas d'oxygène. On les appelle fibres blanches.
- Les unités motrices rapides résistantes à la fatigue stimulent les groupes musculaires de taille moyenne qui ne réagissent pas aussi vite que les unités motrices FF, mais peuvent être maintenues beaucoup plus longtemps (comme le suggère leur nom) et fournissent plus de force que les unités motrices S. Celles-ci utilisent à la fois des moyens oxydatifs et glycolytiques pour gagner de l'énergie.
En plus de la contraction volontaire des muscles squelettiques, les neurones moteurs alpha contribuent également au tonus musculaire , la force continue générée par le muscle non contracté pour s'opposer à l'étirement. Lorsqu'un muscle est étiré, les neurones sensoriels du fuseau neuromusculaire détectent le degré d'étirement et envoient un signal au système nerveux central. Le système nerveux central active les neurones moteurs alpha de la moelle épinière, qui provoquent la contraction des fibres musculaires extrafusales et résistent ainsi à un étirement supplémentaire. Ce processus est également appelé réflexe d'étirement .
- Les motoneurones bêta innervent les fibres musculaires intrafusales des fuseaux musculaires , avec des collatéraux aux fibres extrafusales. Il existe deux types de motoneurones bêta : à contraction lente - Ceux-ci innervent les fibres extrafusales. À contraction rapide - Ceux-ci innervent les fibres intrafusales.
- Les neurones moteurs gamma innervent les fibres musculaires intrafusales présentes dans le fuseau musculaire. Ils régulent la sensibilité du fuseau à l'étirement musculaire. Avec l'activation des neurones gamma, les fibres musculaires intrafusales se contractent de sorte qu'un petit étirement suffit à activer les neurones sensoriels du fuseau et le réflexe d'étirement. Il existe deux types de neurones moteurs gamma : Dynamiques - Ils se concentrent sur les fibres Bag1 et améliorent la sensibilité dynamique. Statiques - Ils se concentrent sur les fibres Bag2 et améliorent la sensibilité à l'étirement.
- Facteurs régulateurs des motoneurones inférieurs
- Principe de taille – il s’agit du soma du motoneurone. Il restreint les neurones plus gros à recevoir un signal d’excitation plus important afin de stimuler les fibres musculaires qu’il innerve. En réduisant le recrutement inutile de fibres musculaires, le corps est capable d’optimiser la consommation d’énergie.
- Courant entrant persistant (PIC) – des études récentes sur des animaux ont montré que le flux constant d’ions tels que le calcium et le sodium à travers les canaux du soma et des dendrites influence l’entrée synaptique. Une autre façon de voir cela est que le neurone post-synaptique est préparé avant de recevoir une impulsion.
- Après l'hyperpolarisation (AHP) – Une tendance a été identifiée qui montre que les motoneurones lents ont des AHP plus intenses pendant une durée plus longue. Une façon de s'en souvenir est que les fibres musculaires lentes peuvent se contracter plus longtemps, il est donc logique que leurs motoneurones correspondants se déclenchent à un rythme plus lent.
Neurones moteurs viscéraux spéciaux
On les appelle également neurones moteurs branchiaux , qui interviennent dans l'expression faciale, la mastication, la phonation et la déglutition. Les nerfs crâniens associés sont les nerfs oculomoteur , abducens , trochléaire et hypoglosse .
Neurones moteurs viscéraux généraux
Ces motoneurones innervent indirectement le muscle cardiaque et les muscles lisses des viscères (les muscles des artères ) : ils font synapse sur des neurones situés dans les ganglions du système nerveux autonome ( sympathique et parasympathique ), localisés dans le système nerveux périphérique (SNP), qui eux-mêmes innervent directement les muscles viscéraux (et aussi certaines cellules glandulaires).
En conséquence, la commande motrice des muscles squelettiques et branchiaux est monosynaptique et n'implique qu'un seul motoneurone, somatique ou branchial , qui fait synapse avec le muscle. En comparaison, la commande motrice des muscles viscéraux est disynaptique et implique deux neurones : le motoneurone viscéral général , situé dans le SNC, fait synapse avec un neurone ganglionnaire, situé dans le SNP, qui fait synapse avec le muscle.
Tous les motoneurones des vertébrés sont cholinergiques , c'est-à-dire qu'ils libèrent le neurotransmetteur acétylcholine . Les neurones ganglionnaires parasympathiques sont également cholinergiques, tandis que la plupart des neurones ganglionnaires sympathiques sont noradrénergiques , c'est-à-dire qu'ils libèrent le neurotransmetteur noradrénaline . (voir tableau)
Jonctions neuromusculaires
Un seul motoneurone peut innerver de nombreuses fibres musculaires et une fibre musculaire peut subir de nombreux potentiels d'action dans le temps nécessaire à une seule secousse musculaire . En conséquence, si un potentiel d'action arrive avant qu'une secousse ne soit terminée, les secousses peuvent se superposer les unes aux autres, soit par sommation , soit par une contraction tétanique . En somme, le muscle est stimulé de manière répétitive de telle sorte que des potentiels d'action supplémentaires provenant du système nerveux somatique arrivent avant la fin de la secousse. Les secousses se superposent ainsi les unes aux autres, ce qui entraîne une force supérieure à celle d'une seule secousse. Une contraction tétanique est provoquée par une stimulation constante à très haute fréquence - les potentiels d'action arrivent à un rythme si rapide que les secousses individuelles sont indiscernables et la tension augmente progressivement pour finalement atteindre un plateau.
L'interface entre un motoneurone et une fibre musculaire est une synapse spécialisée appelée jonction neuromusculaire . Lors d'une stimulation adéquate, le motoneurone libère un flot de neurotransmetteurs acétylcholine (Ach) à partir de vésicules synaptiques liées à la membrane plasmique des terminaisons axonales. Les molécules d'acétylcholine se lient aux récepteurs postsynaptiques présents dans la plaque terminale motrice. Une fois que deux récepteurs d'acétylcholine sont liés, un canal ionique est ouvert et les ions sodium peuvent pénétrer dans la cellule. L'afflux de sodium dans la cellule provoque une dépolarisation et déclenche un potentiel d'action musculaire. Les tubules T du sarcolemme sont ensuite stimulés pour provoquer la libération d'ions calcium à partir du réticulum sarcoplasmique. C'est cette libération chimique qui provoque la contraction de la fibre musculaire cible.
Chez les invertébrés , selon le neurotransmetteur libéré et le type de récepteur auquel il se lie, la réponse de la fibre musculaire peut être soit excitatrice, soit inhibitrice. Chez les vertébrés , en revanche, la réponse d'une fibre musculaire à un neurotransmetteur ne peut être qu'excitatrice, c'est-à-dire contractile. La relaxation musculaire et l'inhibition de la contraction musculaire chez les vertébrés ne s'obtiennent que par inhibition du motoneurone lui-même. C'est ainsi que les myorelaxants agissent en agissant sur les motoneurones qui innervent les muscles (en diminuant leur activité électrophysiologique ) ou sur les jonctions neuromusculaires cholinergiques , plutôt que sur les muscles eux-mêmes.
Entrée synaptique vers les motoneurones
Les neurones moteurs reçoivent des entrées synaptiques des neurones prémoteurs. Les neurones prémoteurs peuvent être 1) des interneurones spinaux qui ont des corps cellulaires dans la moelle épinière, 2) des neurones sensoriels qui transmettent des informations de la périphérie et des synapses directement sur les motoneurones , 3) des neurones descendants qui transmettent des informations du cerveau et du tronc cérébral . Les synapses peuvent être excitatrices , inhibitrices , électriques ou neuromodulatrices . Pour un neurone moteur donné, il est difficile de déterminer la contribution relative des différentes sources d'entrée, mais les progrès de la connectomique ont rendu cela possible pour les neurones moteurs de la mouche à fruits . Chez la mouche, les neurones moteurs contrôlant les pattes et les ailes se trouvent dans la moelle nerveuse ventrale , homologue de la moelle épinière . Les neurones moteurs de la mouche varient de plus de 100X dans le nombre total de synapses d'entrée. Cependant, chaque motoneurone reçoit des fractions similaires de ses synapses de chaque source prémotrice : environ 70 % des neurones du VNC, environ 10 % des neurones descendants, environ 3 % des neurones sensoriels et environ 6 % des neurones du VNC qui envoient également un processus vers le cerveau. Les 10 % restants des synapses proviennent de fragments neuronaux qui ne sont pas identifiés par les algorithmes de segmentation d'images actuels et nécessitent une segmentation manuelle supplémentaire pour être mesurés.