L' enveloppe nucléaire , également connue sous le nom de membrane nucléaire , est constituée de deux membranes bicouches lipidiques qui, dans les cellules eucaryotes, entourent le noyau , qui renferme le matériel génétique .
L'enveloppe nucléaire est constituée de deux membranes bicouches lipidiques : une membrane nucléaire interne et une membrane nucléaire externe. L'espace entre les membranes est appelé espace périnucléaire. Il mesure généralement entre 10 et 50 nm de large. La membrane nucléaire externe est continue avec la membrane du réticulum endoplasmique . L'enveloppe nucléaire possède de nombreux pores nucléaires qui permettent aux matériaux de se déplacer entre le cytosol et le noyau. Des protéines filamentaires intermédiaires appelées lamines forment une structure appelée lamina nucléaire sur la face interne de la membrane nucléaire interne et fournissent un support structurel au noyau.
Structure
L'enveloppe nucléaire est constituée de deux membranes bicouches lipidiques, une membrane nucléaire interne et une membrane nucléaire externe. Ces membranes sont reliées entre elles par des pores nucléaires. Deux ensembles de filaments intermédiaires assurent le support de l'enveloppe nucléaire. Un réseau interne forme la lame nucléaire sur la membrane nucléaire interne. Un réseau plus lâche se forme à l'extérieur pour fournir un support externe. La forme réelle de l'enveloppe nucléaire est irrégulière. Elle présente des invaginations et des protubérances et peut être observée au microscope électronique .
Membrane externe
La membrane nucléaire externe partage également une frontière commune avec le réticulum endoplasmique . Bien qu'elle soit physiquement liée, la membrane nucléaire externe contient des protéines trouvées en concentrations bien plus élevées que le réticulum endoplasmique. Les quatre protéines de nesprine ( protéines de répétition de la spectrine de l'enveloppe nucléaire ) présentes chez les mammifères sont exprimées dans la membrane nucléaire externe. Les protéines de nesprine relient les filaments du cytosquelette au nucléosquelette. Les connexions médiées par la nesprine au cytosquelette contribuent au positionnement nucléaire et à la fonction mécanosensorielle de la cellule. du domaine KASH de Nesprin-1 et -2 font partie d'un complexe LINC (lieur du nucléosquelette et du cytosquelette) et peuvent se lier directement aux composants du cystosquelette, tels que les filaments d'actine , ou peuvent se lier aux protéines de l'espace périnucléaire. Les protéines Nesprin-3 et -4 peuvent jouer un rôle dans le déchargement d'une énorme cargaison ; les protéines Nesprin-3 se lient à la plectine et lient l'enveloppe nucléaire aux filaments intermédiaires cytoplasmiques. Les protéines Nesprin-4 se lient à la kinésine-1 motrice dirigée vers l'extrémité positive. La membrane nucléaire externe est également impliquée dans le développement, car elle fusionne avec la membrane nucléaire interne pour former des pores nucléaires.
Membrane interne
La membrane nucléaire interne entoure le nucléoplasme et est recouverte par la lame nucléaire , un maillage de filaments intermédiaires qui stabilise la membrane nucléaire et participe à la fonction de la chromatine . Elle est reliée à la membrane externe par des pores nucléaires qui pénètrent dans les membranes. Bien que les deux membranes et le réticulum endoplasmique soient liés, les protéines intégrées dans les membranes ont tendance à rester en place plutôt que de se disperser à travers le continuum. Elle est tapissée d'un réseau de fibres appelé la lame nucléaire qui a une épaisseur de 10 à 40 nm et assure la résistance.
Des mutations dans les gènes codant pour les protéines de la membrane nucléaire interne peuvent provoquer plusieurs laminopathies .
Pores nucléaires
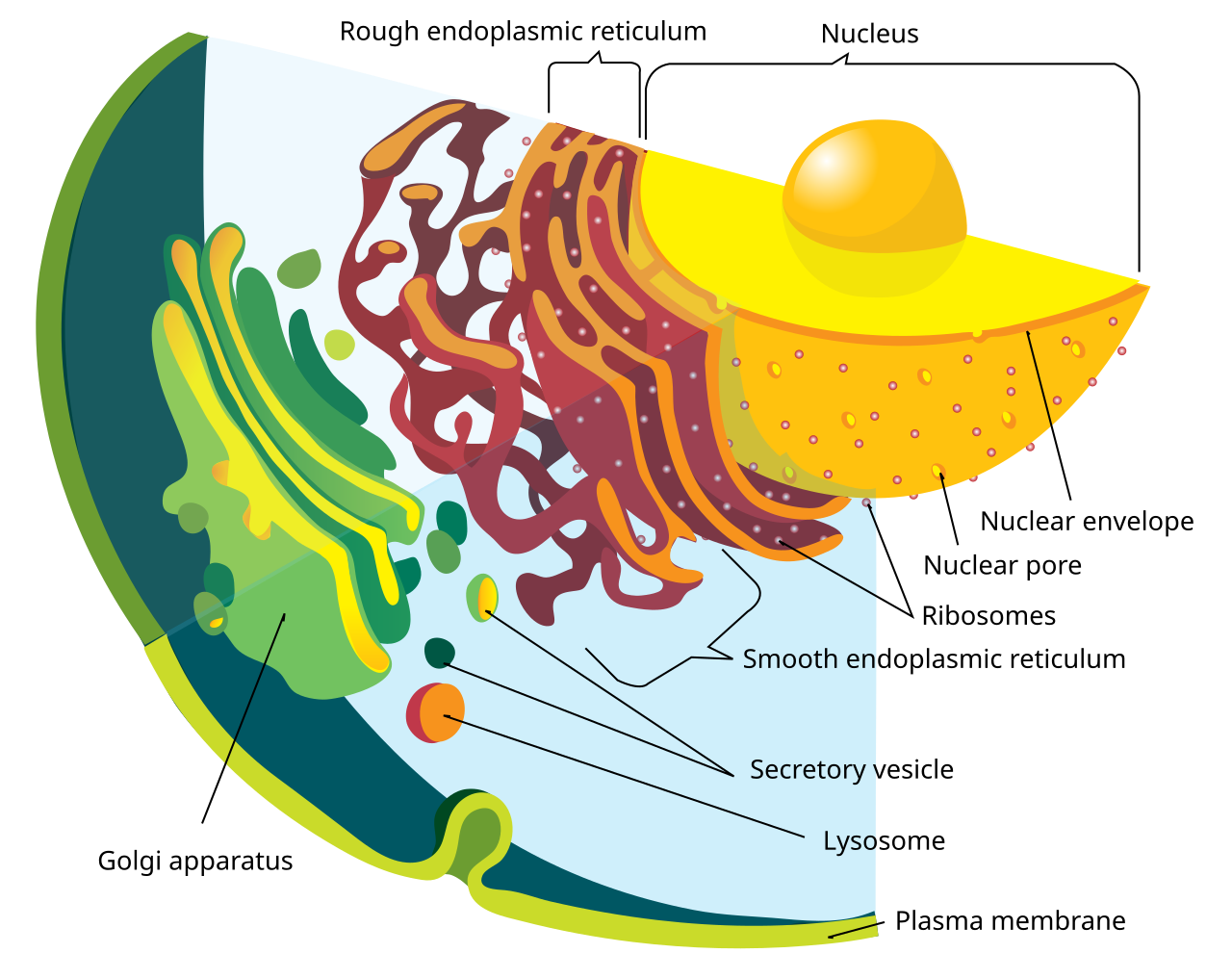
L'enveloppe nucléaire est percée par environ un millier de complexes de pores nucléaires , d'environ 100 nm de diamètre, avec un canal interne d'environ 40 nm de large. Les complexes contiennent un certain nombre de nucléoporines , des protéines qui relient les membranes nucléaires internes et externes.
Division cellulaire
Au cours de la phase G2 de l'interphase , la membrane nucléaire augmente sa surface et double le nombre de ses complexes de pores nucléaires. Chez les eucaryotes comme la levure qui subissent une mitose fermée , la membrane nucléaire reste intacte pendant la division cellulaire. Les fibres du fuseau se forment à l'intérieur de la membrane ou la pénètrent sans la déchirer. Chez d'autres eucaryotes (animaux comme plantes), la membrane nucléaire doit se décomposer au cours de la phase prométaphase de la mitose pour permettre aux fibres du fuseau mitotique d'accéder aux chromosomes à l'intérieur. Les processus de décomposition et de reformation ne sont pas bien compris.
Panne
Chez les mammifères, la membrane nucléaire peut se décomposer en quelques minutes, en suivant une série d'étapes au cours des premiers stades de la mitose . Tout d'abord, les M-Cdk phosphorylent les polypeptides de nucléoporine et ils sont sélectivement retirés des complexes de pores nucléaires. Après cela, le reste des complexes de pores nucléaires se décompose simultanément. Des preuves biochimiques suggèrent que les complexes de pores nucléaires se désassemblent en morceaux stables plutôt que de se désintégrer en petits fragments polypeptidiques. Les M-Cdk phosphorylent également des éléments de la lame nucléaire (la structure qui soutient l'enveloppe) conduisant au désassemblage de la lame et donc des membranes de l'enveloppe en petites vésicules. La microscopie électronique et à fluorescence a fourni des preuves solides que la membrane nucléaire est absorbée par le réticulum endoplasmique - des protéines nucléaires qui ne se trouvent normalement pas dans le réticulum endoplasmique apparaissent pendant la mitose.
En plus de la rupture de la membrane nucléaire au cours de la phase prométaphase de la mitose , la membrane nucléaire se rompt également dans les cellules de mammifères en migration au cours de la phase interphasique du cycle cellulaire. Cette rupture transitoire est probablement causée par une déformation nucléaire. La rupture est rapidement réparée par un processus dépendant des « complexes de tri endosomal requis pour le transport » ( ESCRT ) constitués de complexes protéiques cytosoliques . Au cours des événements de rupture de la membrane nucléaire, des cassures double brin d'ADN se produisent. Ainsi, la survie des cellules migrant dans des environnements confinés semble dépendre de l'efficacité de l'enveloppe nucléaire et des mécanismes de réparation de l'ADN .
Une dégradation aberrante de l'enveloppe nucléaire a également été observée dans les laminopathies et dans les cellules cancéreuses, entraînant une mauvaise localisation des protéines cellulaires, la formation de micronoyaux et une instabilité génomique.
Réformation
La manière exacte dont la membrane nucléaire se reforme pendant la télophase de la mitose est sujette à débat. Deux théories existent —
- Fusion des vésicules : les vésicules de la membrane nucléaire fusionnent pour reconstruire la membrane nucléaire
- Remodelage du réticulum endoplasmique : où les parties du réticulum endoplasmique contenant la membrane nucléaire absorbée enveloppent l’espace nucléaire, reformant une membrane fermée.
Origine de la membrane nucléaire
Une étude comparative de la génomique , de l'évolution et des origines de la membrane nucléaire a conduit à l'hypothèse selon laquelle le noyau aurait émergé chez l' ancêtre eucaryote primitif (le « précaryote ») et aurait été déclenché par la symbiose archéobactérienne . Plusieurs idées ont été proposées pour l'origine évolutive de la membrane nucléaire. Ces idées incluent l'invagination de la membrane plasmique chez un ancêtre procaryote, ou la formation d'un véritable nouveau système membranaire suite à l'établissement de proto- mitochondries dans l'hôte archéen. La fonction adaptative de la membrane nucléaire pourrait avoir été de servir de barrière pour protéger le génome des espèces réactives de l'oxygène (ROS) produites par les pré-mitochondries des cellules.