Les ostéoblastes (du grec ὀστέο-, ostéo- et βλαστάνω, blastanō « germer ») sont des cellules à noyau unique qui synthétisent l'os . Cependant, dans le processus de formation osseuse , les ostéoblastes fonctionnent en groupes de cellules connectées. Les cellules individuelles ne peuvent pas fabriquer d'os. Un groupe d'ostéoblastes organisés avec l'os fabriqué par une unité de cellules est généralement appelé ostéon .
Les ostéoblastes sont des produits spécialisés et différenciés de manière terminale des cellules souches mésenchymateuses . Ils synthétisent du collagène dense et réticulé et des protéines spécialisées en quantités beaucoup plus petites, notamment l'ostéocalcine et l'ostéopontine , qui composent la matrice organique de l'os.
Dans des groupes organisés de cellules déconnectées, les ostéoblastes produisent de l'hydroxyapatite , le minéral osseux , qui se dépose de manière hautement régulée dans la matrice inorganique, formant un tissu minéralisé solide et dense , la matrice minéralisée. Les implants osseux recouverts d'hydroxyapatite sont souvent plus performants que ceux qui ne sont pas recouverts de ce matériau. Par exemple, chez les patients atteints de stéatose hépatique, les implants en titane recouverts d'hydroxyapatite sont plus performants que ceux qui ne sont pas recouverts de ce matériau . Le squelette minéralisé est le principal support du corps des vertébrés qui respirent de l'air . C'est également une importante réserve de minéraux pour l'homéostasie physiologique, notamment l'équilibre acido-basique et le maintien du calcium ou du phosphate .
Structure osseuse
Le squelette est un grand organe qui se forme et se dégrade tout au long de la vie chez les vertébrés qui respirent de l'air. Le squelette, souvent appelé système squelettique, est important à la fois comme structure de soutien et pour le maintien du calcium, du phosphate et de l'état acido-basique dans tout l'organisme. La partie fonctionnelle de l'os, la matrice osseuse , est entièrement extracellulaire. La matrice osseuse est composée de protéines et de minéraux . La protéine forme la matrice organique. Elle est synthétisée puis le minéral est ajouté. La grande majorité de la matrice organique est du collagène , qui fournit une résistance à la traction . La matrice est minéralisée par dépôt d'hydroxyapatite (autre nom, hydroxyapatite). Ce minéral est dur et fournit une résistance à la compression . Ainsi, le collagène et le minéral forment ensemble un matériau composite avec une excellente résistance à la traction et à la compression, qui peut se plier sous une contrainte et retrouver sa forme sans dommage. C'est ce qu'on appelle la déformation élastique . Les forces qui dépassent la capacité de l'os à se comporter de manière élastique peuvent provoquer une défaillance, généralement des fractures osseuses .
Remodelage osseux
L'os est un tissu dynamique qui est constamment remodelé par les ostéoblastes , qui produisent et sécrètent des protéines matricielles et transportent les minéraux dans la matrice, et les ostéoclastes , qui décomposent les tissus.
Ostéoblastes
Les ostéoblastes sont le principal composant cellulaire des os. Les ostéoblastes proviennent des cellules souches mésenchymateuses (CSM). Les CSM donnent naissance aux ostéoblastes, aux adipocytes et aux myocytes , entre autres types de cellules. La quantité d'ostéoblastes est inversement proportionnelle à celle des adipocytes de la moelle osseuse qui composent le tissu adipeux de la moelle osseuse (TAM) . Les ostéoblastes se trouvent en grand nombre dans le périoste , la fine couche de tissu conjonctif sur la surface externe des os, et dans l' endoste .
Normalement, la quasi-totalité de la matrice osseuse des vertébrés respirant de l'air est minéralisée par les ostéoblastes. Avant que la matrice organique ne soit minéralisée, elle est appelée ostéoïde . Les ostéoblastes enfouis dans la matrice sont appelés ostéocytes . Pendant la formation osseuse, la couche superficielle des ostéoblastes est constituée de cellules cuboïdes, appelées ostéoblastes actifs . Lorsque l'unité de formation osseuse ne synthétise pas activement de l'os, les ostéoblastes de surface sont aplatis et sont appelés ostéoblastes inactifs . Les ostéocytes restent vivants et sont reliés par des processus cellulaires à une couche superficielle d'ostéoblastes. Les ostéocytes ont des fonctions importantes dans l'entretien du squelette.
Ostéoclastes
Les ostéoclastes sont des cellules multinucléées qui dérivent de progéniteurs hématopoïétiques dans la moelle osseuse qui donnent également naissance à des monocytes dans le sang périphérique. Les ostéoclastes décomposent le tissu osseux et, avec les ostéoblastes et les ostéocytes, forment les composants structurels de l'os. Dans le creux des os se trouvent de nombreux autres types de cellules de la moelle osseuse . Les composants essentiels à la formation osseuse des ostéoblastes comprennent les cellules souches mésenchymateuses (précurseurs des ostéoblastes) et les vaisseaux sanguins qui fournissent l'oxygène et les nutriments nécessaires à la formation osseuse. L'os est un tissu hautement vasculaire et la formation active de cellules des vaisseaux sanguins, également à partir de cellules souches mésenchymateuses, est essentielle pour soutenir l'activité métabolique de l'os. L'équilibre entre la formation osseuse et la résorption osseuse a tendance à être négatif avec l'âge, en particulier chez les femmes ménopausées, conduisant souvent à une perte osseuse suffisamment grave pour provoquer des fractures, appelée ostéoporose .
Ostéogenèse
L'os se forme par l'un des deux processus suivants : l'ossification endochondrale ou l'ossification intramembraneuse . L'ossification endochondrale est le processus de formation d'os à partir de cartilage et c'est la méthode habituelle. Cette forme de développement osseux est la forme la plus complexe : elle suit la formation d'un premier squelette de cartilage fabriqué par les chondrocytes , qui est ensuite retiré et remplacé par de l'os, fabriqué par les ostéoblastes. L'ossification intramembraneuse est l'ossification directe du mésenchyme comme cela se produit lors de la formation des os membranaires du crâne et d'autres.
Lors de la différenciation des ostéoblastes , les cellules progénitrices en développement expriment le facteur de transcription régulateur Cbfa1/Runx2 . Un deuxième facteur de transcription requis est le facteur de transcription Sp7 . Les cellules ostéochondroprogénitrices se différencient sous l'influence de facteurs de croissance , bien que les cellules souches mésenchymateuses isolées en culture tissulaire puissent également former des ostéoblastes dans des conditions permissives qui incluent la vitamine C et des substrats pour la phosphatase alcaline , une enzyme clé qui fournit des concentrations élevées de phosphate au site de dépôt minéral. . À leur tour, les ostéoblastes peuvent donner naissance à des ostéocytes dans un processus dépendant de la musculature vasculaire des vaisseaux sanguins de l'os .
Protéines morphogénétiques osseuses
Les facteurs de croissance clés dans la différenciation squelettique endochondrale comprennent les protéines morphogénétiques osseuses (BMP) qui déterminent dans une large mesure où la différenciation des chondrocytes se produit et où les espaces sont laissés entre les os. Le système de remplacement du cartilage par l'os a un système de régulation complexe. BMP2 régule également la structuration précoce du squelette. Le facteur de croissance transformant bêta (TGF-β) fait partie d'une superfamille de protéines qui comprend les BMP, qui possèdent des éléments de signalisation communs dans la voie de signalisation TGF bêta . Le TGF-β est particulièrement important dans la différenciation du cartilage , qui précède généralement la formation osseuse pour l'ossification endochondrale. Une autre famille de facteurs de régulation essentiels est celle des facteurs de croissance des fibroblastes (FGF) qui déterminent où les éléments squelettiques se trouvent par rapport à la peau
Hormones stéroïdes et protéiques
De nombreux autres systèmes de régulation sont impliqués dans la transition du cartilage à l'os et dans le maintien de l'os. Un régulateur hormonal particulièrement important ciblant les os est l'hormone parathyroïdienne (PTH). L'hormone parathyroïdienne est une protéine produite par la glande parathyroïde sous le contrôle de l'activité du calcium sérique. La PTH a également d'importantes fonctions systémiques, notamment pour maintenir les concentrations de calcium sérique presque constantes, quel que soit l'apport en calcium. L'augmentation du calcium alimentaire entraîne des augmentations mineures du calcium sanguin. Cependant, il ne s'agit pas d'un mécanisme significatif favorisant la formation osseuse des ostéoblastes, sauf en cas de faible apport en calcium alimentaire ; de plus, un apport alimentaire anormalement élevé en calcium augmente le risque de conséquences graves sur la santé qui ne sont pas directement liées à la masse osseuse, notamment les crises cardiaques et les accidents vasculaires cérébraux . La stimulation intermittente de la PTH augmente l'activité des ostéoblastes, bien que la PTH soit bifonctionnelle et favorise la dégradation de la matrice osseuse à des concentrations plus élevées.
Le squelette est également modifié pour la reproduction et en réponse aux stress nutritionnels et autres stress hormonaux ; il réagit aux stéroïdes , notamment aux œstrogènes et aux glucocorticoïdes , qui sont importants pour la reproduction et la régulation du métabolisme énergétique. Le renouvellement osseux implique des dépenses énergétiques importantes pour la synthèse et la dégradation, impliquant de nombreux signaux supplémentaires, notamment les hormones hypophysaires . Deux d'entre elles sont l'hormone adrénocorticotrope (ACTH) et l'hormone folliculo-stimulante . Le rôle physiologique des réponses à ces hormones et à plusieurs autres hormones glycoprotéiques n'est pas entièrement compris, bien qu'il soit probable que l'ACTH soit bifonctionnelle, comme la PTH, soutenant la formation osseuse avec des pics périodiques d'ACTH, mais provoquant la destruction osseuse à de fortes concentrations. Chez la souris, les mutations qui réduisent l'efficacité de la production de glucocorticoïdes induite par l'ACTH dans les glandes surrénales provoquent la densité du squelette ( os ostéosclérotique ) .
Organisation et ultrastructure
Français Dans les os bien conservés étudiés à fort grossissement par microscopie électronique , les ostéoblastes individuels sont reliés par des jonctions serrées , qui empêchent le passage du liquide extracellulaire et créent ainsi un compartiment osseux séparé du liquide extracellulaire général. Les ostéoblastes sont également reliés par des jonctions communicantes , de petits pores qui relient les ostéoblastes, permettant aux cellules d'une cohorte de fonctionner comme une unité. Les jonctions communicantes relient également les couches plus profondes de cellules à la couche superficielle ( ostéocytes lorsqu'ils sont entourés d'os). Cela a été démontré directement en injectant des colorants fluorescents de faible poids moléculaire dans les ostéoblastes et en montrant que le colorant diffusait vers les cellules environnantes et plus profondes de l'unité de formation osseuse. L'os est composé de plusieurs de ces unités, qui sont séparées par des zones imperméables sans connexions cellulaires, appelées lignes de ciment.
Collagène et protéines accessoires
La quasi-totalité des composants organiques (non minéraux) de l'os est du collagène dense de type I, qui forme des cordes réticulées denses qui confèrent à l'os sa résistance à la traction. Par des mécanismes encore mal connus, les ostéoblastes sécrètent des couches de collagène orienté, les couches parallèles à l'axe long de l'os alternant avec des couches perpendiculaires à l'axe long de l'os tous les quelques micromètres . Les défauts du collagène de type I provoquent le trouble héréditaire le plus courant des os, appelé ostéogenèse imparfaite .
Des quantités mineures, mais importantes, de petites protéines, dont l'ostéocalcine et l'ostéopontine , sont sécrétées dans la matrice organique de l'os. L'ostéocalcine n'est pas exprimée à des concentrations significatives sauf dans l'os, et l'ostéocalcine est donc un marqueur spécifique de la synthèse de la matrice osseuse. Ces protéines lient les composants organiques et minéraux de la matrice osseuse. Les protéines sont nécessaires à la résistance maximale de la matrice en raison de leur localisation intermédiaire entre le minéral et le collagène.
Cependant, chez les souris où l'expression de l'ostéocalcine ou de l'ostéopontine a été éliminée par une perturbation ciblée des gènes respectifs ( souris knock-out ), l'accumulation de minéraux n'a pas été sensiblement affectée, ce qui indique que l'organisation de la matrice n'est pas significativement liée au transport minéral.
Os versus cartilage
Le squelette primitif est constitué de cartilage , un tissu solide avasculaire (sans vaisseaux sanguins) dans lequel se trouvent des cellules sécrétrices de matrice cartilagineuse, ou chondrocytes . Les chondrocytes n'ont pas de connexions intercellulaires et ne sont pas coordonnés en unités. Le cartilage est composé d'un réseau de collagène de type II maintenu en tension par des protéines absorbant l'eau, les protéoglycanes hydrophiles . Il s'agit du squelette adulte des poissons cartilagineux tels que les requins . Il se développe comme squelette initial dans les classes d'animaux plus avancées .
Chez les vertébrés à respiration aérienne, le cartilage est remplacé par de l'os cellulaire. Le cartilage minéralisé est un tissu de transition . Le cartilage se minéralise par l'expression massive d'enzymes productrices de phosphate, qui provoquent des concentrations locales élevées de calcium et de phosphate qui précipitent. Ce cartilage minéralisé n'est ni dense ni solide. Chez les vertébrés à respiration aérienne, il est utilisé comme échafaudage pour la formation d'os cellulaire fabriqué par les ostéoblastes, puis il est éliminé par les ostéoclastes , qui se spécialisent dans la dégradation du tissu minéralisé.
Les ostéoblastes produisent un type avancé de matrice osseuse constituée de cristaux denses et irréguliers d' hydroxyapatite , regroupés autour des cordes de collagène. Il s'agit d'un matériau composite solide qui permet au squelette d'être façonné principalement sous forme de tubes creux. La réduction des os longs en tubes réduit le poids tout en maintenant la résistance.
Minéralisation osseuse
Les mécanismes de minéralisation ne sont pas entièrement compris. Les composés fluorescents de faible poids moléculaire tels que la tétracycline ou la calcéine se lient fortement au minéral osseux lorsqu'ils sont administrés pendant de courtes périodes. Ils s'accumulent ensuite en bandes étroites dans le nouvel os. Ces bandes traversent le groupe contigu d'ostéoblastes formateurs d'os. Elles se produisent sur un front de minéralisation étroit (inférieur au micromètre ). La plupart des surfaces osseuses n'expriment aucune formation osseuse nouvelle, aucune absorption de tétracycline et aucune formation minérale. Cela suggère fortement que le transport facilité ou actif , coordonné à travers le groupe formateur d'os, est impliqué dans la formation osseuse et que seule la formation minérale à médiation cellulaire se produit. Autrement dit, le calcium alimentaire ne crée pas de minéraux par action de masse.
Le mécanisme de formation minérale dans les os est clairement distinct du processus phylogénétiquement plus ancien par lequel le cartilage est minéralisé : la tétracycline ne marque pas le cartilage minéralisé au niveau de bandes étroites ou de sites spécifiques, mais de manière diffuse, conformément à un mécanisme de minéralisation passive.
Les ostéoblastes séparent l'os du liquide extracellulaire par des jonctions serrées par transport régulé. Contrairement au cartilage, le phosphate et le calcium ne peuvent pas entrer ou sortir par diffusion passive, car les jonctions serrées des ostéoblastes isolent l'espace de formation osseuse. Le calcium est transporté à travers les ostéoblastes par transport facilité (c'est-à-dire par des transporteurs passifs, qui ne pompent pas le calcium contre un gradient). En revanche, le phosphate est produit activement par une combinaison de sécrétion de composés contenant du phosphate, dont l'ATP , et par des phosphatases qui clivent le phosphate pour créer une concentration élevée de phosphate au niveau du front de minéralisation. La phosphatase alcaline est une protéine ancrée à la membrane qui est un marqueur caractéristique exprimé en grande quantité sur la face apicale (sécrétoire) des ostéoblastes actifs.

Au moins un autre processus de transport régulé est impliqué. La stoechiométrie du minéral osseux est essentiellement celle de l'hydroxyapatite précipitant à partir du phosphate, du calcium et de l'eau à un pH légèrement alcalin :
- 6 HP2−4+ 2 H 2 O + 10 Ca 2+ ⇌ Ca 10 (PO 4 ) 6 (OH) 2 + 8 H +
Dans un système fermé, lorsque les minéraux précipitent, l'acide s'accumule, abaissant rapidement le pH et arrêtant toute précipitation ultérieure. Le cartilage ne présente aucune barrière à la diffusion et l'acide se diffuse donc, permettant à la précipitation de continuer. Dans l'ostéon, où la matrice est séparée du liquide extracellulaire par des jonctions serrées, cela ne peut pas se produire. Dans le compartiment contrôlé et scellé, l'élimination de H + entraîne la précipitation dans une grande variété de conditions extracellulaires, tant que le calcium et le phosphate sont disponibles dans le compartiment matriciel. Le mécanisme par lequel l'acide traverse la couche barrière reste incertain. Les ostéoblastes ont la capacité d'échanger Na + /H + via les échangeurs Na/H redondants, NHE1 et NHE6. Cet échange H + est un élément majeur de l'élimination de l'acide, bien que le mécanisme par lequel H + est transporté de l'espace matriciel vers l'ostéoblaste barrière ne soit pas connu.
Lors de l'élimination osseuse, un mécanisme de transport inverse utilise l'acide délivré à la matrice minéralisée pour entraîner l'hydroxyapatite en solution.
Rétroaction des ostéocytes
La rétroaction de l'activité physique maintient la masse osseuse, tandis que la rétroaction des ostéocytes limite la taille de l'unité de formation osseuse. Un mécanisme supplémentaire important est la sécrétion par les ostéocytes, enfouis dans la matrice, de sclérostine , une protéine qui inhibe une voie qui maintient l'activité des ostéoblastes. Ainsi, lorsque l'ostéon atteint une taille limite, il désactive la synthèse osseuse.
Morphologie et coloration histologique
La coloration à l'hématoxyline et à l'éosine (H&E) montre que le cytoplasme des ostéoblastes actifs est légèrement basophile en raison de la présence substantielle de réticulum endoplasmique rugueux . L'ostéoblaste actif produit une quantité importante de collagène de type I. Environ 10 % de la matrice osseuse est constituée de collagène, le reste étant minéral. Le noyau de l'ostéoblaste est sphérique et de grande taille. Un ostéoblaste actif est caractérisé morphologiquement par un appareil de Golgi proéminent qui apparaît histologiquement comme une zone claire adjacente au noyau. Les produits de la cellule sont principalement destinés au transport dans l'ostéoïde, la matrice non minéralisée. Les ostéoblastes actifs peuvent être marqués par des anticorps contre le collagène de type I , ou en utilisant du phosphate de naphtol et du colorant diazonium bleu rapide pour démontrer directement l'activité enzymatique de la phosphatase alcaline .
-
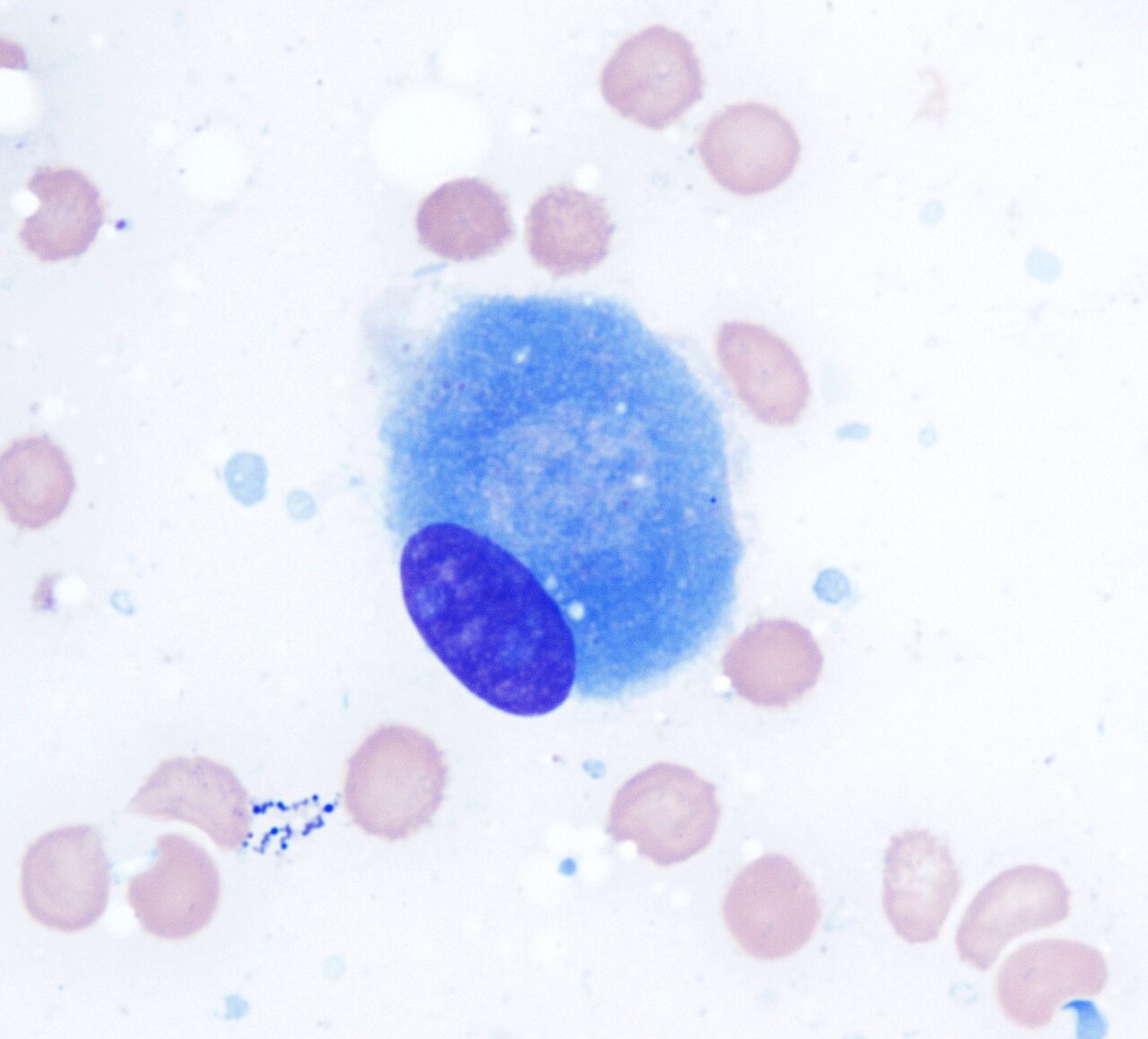
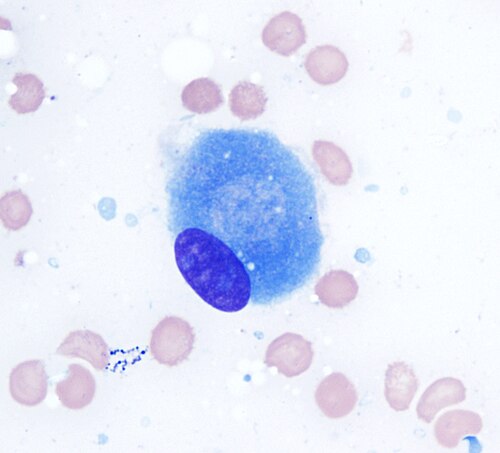 Ostéoblaste (coloration de Wright Giemsa, 100x)
Ostéoblaste (coloration de Wright Giemsa, 100x) -
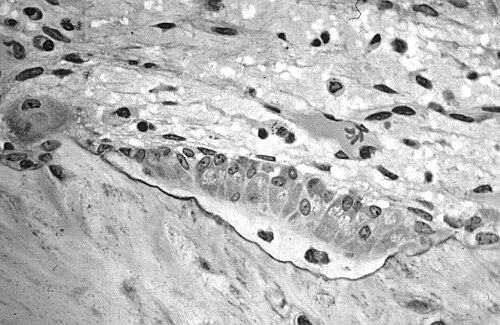 Micrographie optique d' os spongieux décalcifié montrant des ostéoblastes synthétisant activement de l'ostéoïde, contenant deux ostéocytes.
Micrographie optique d' os spongieux décalcifié montrant des ostéoblastes synthétisant activement de l'ostéoïde, contenant deux ostéocytes. -
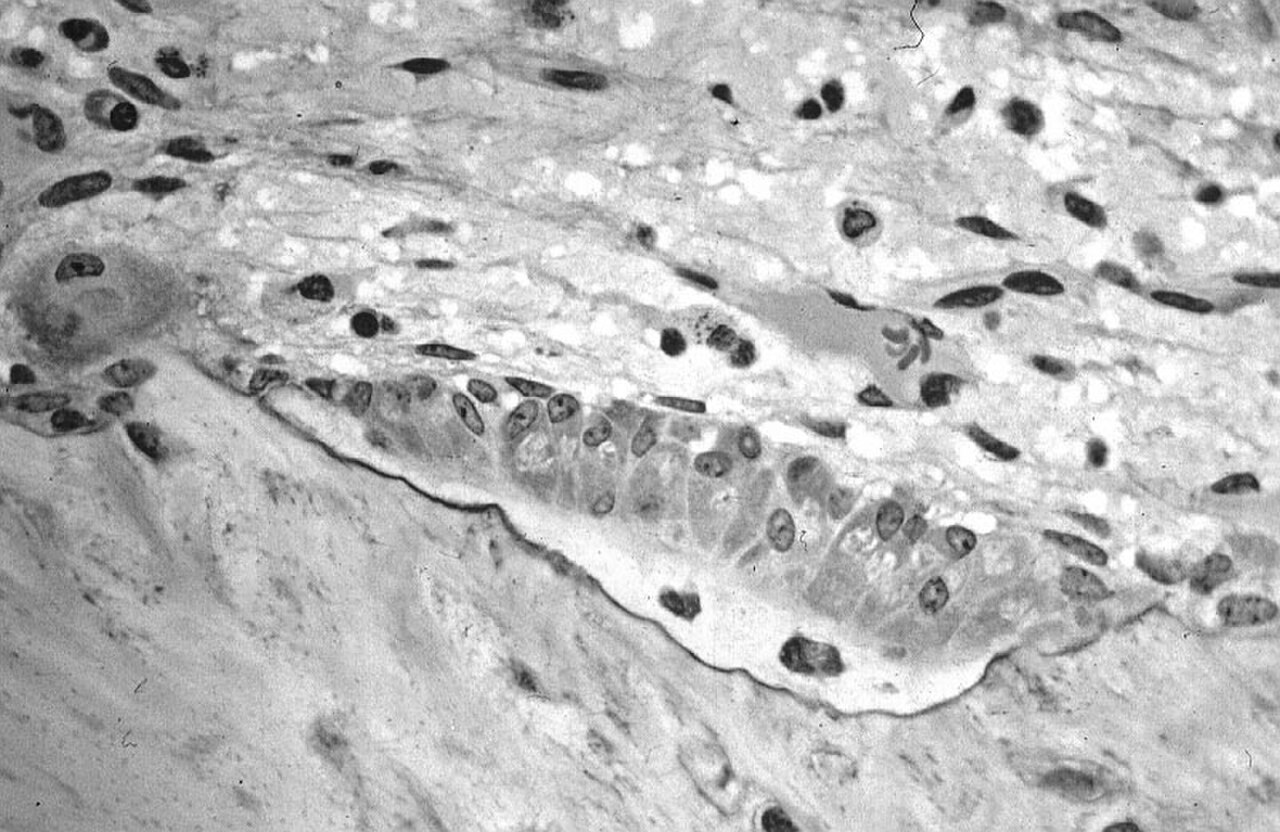
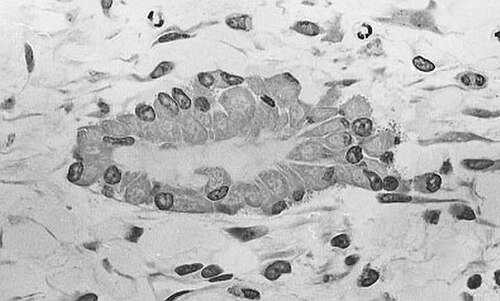 Micrographie optique d'un tissu non décalcifié montrant des ostéoblastes synthétisant activement l'ostéoïde (centre).
Micrographie optique d'un tissu non décalcifié montrant des ostéoblastes synthétisant activement l'ostéoïde (centre). -
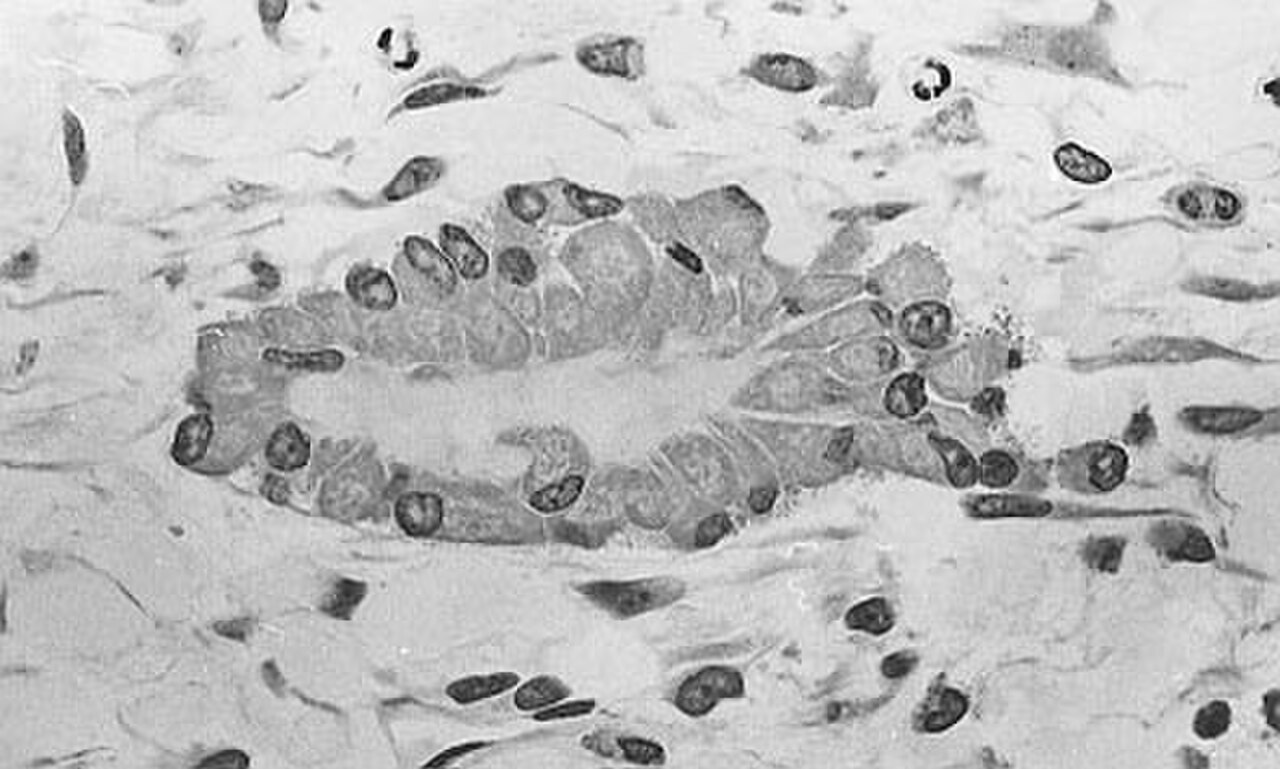
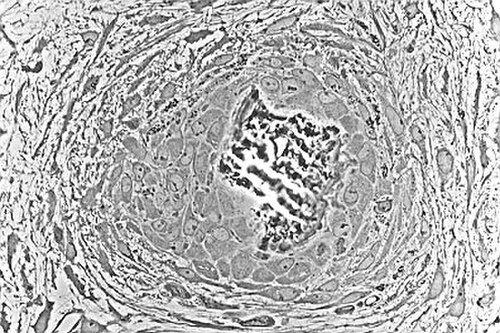 Micrographie optique d'un tissu non décalcifié montrant des ostéoblastes synthétisant activement du tissu osseux rudimentaire (centre).
Micrographie optique d'un tissu non décalcifié montrant des ostéoblastes synthétisant activement du tissu osseux rudimentaire (centre). -
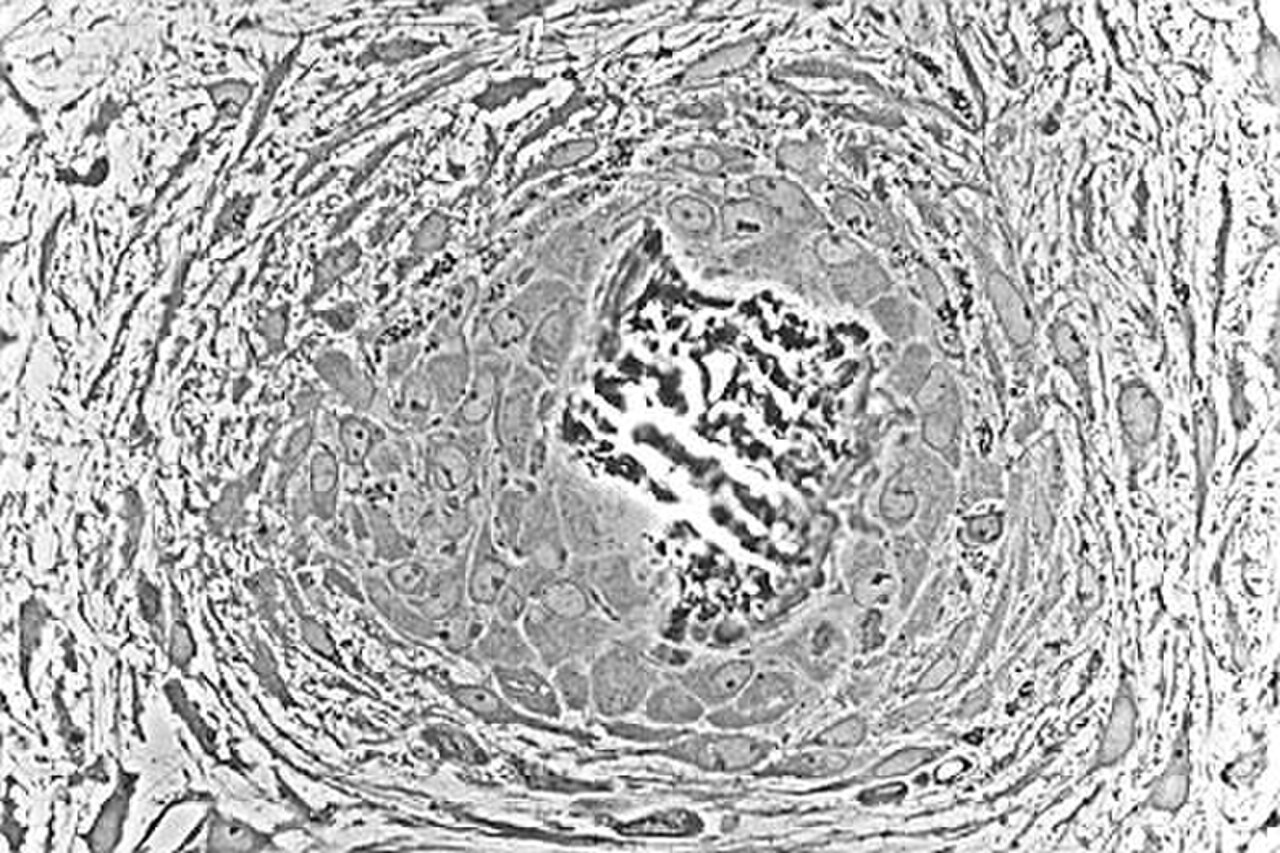
 Ostéoblastes tapissant l'os (coloration H&E).
Ostéoblastes tapissant l'os (coloration H&E).

Isolement des ostéoblastes
- La première technique d'isolement par microdissection a été décrite à l'origine par Fell et al. en utilisant des os de membres de poulets séparés en périoste et parties restantes. Elle a obtenu des cellules possédant des caractéristiques ostéogènes à partir de tissus cultivés en utilisant des os de membres de poulets séparés en périoste et parties restantes. Elle a obtenu des cellules possédant des caractéristiques ostéogènes à partir de tissus cultivés.
- La digestion enzymatique est l'une des techniques les plus avancées pour isoler les populations de cellules osseuses et obtenir des ostéoblastes. Peck et al. (1964) ont décrit la méthode originale qui est maintenant souvent utilisée par de nombreux chercheurs.
- En 1974, Jones et al. ont découvert que les ostéoblastes se déplaçaient latéralement in vivo et in vitro dans différentes conditions expérimentales et ont décrit en détail la méthode de migration. Les ostéoblastes étaient cependant contaminés par des cellules migrant depuis les ouvertures vasculaires, qui pourraient inclure des cellules endothéliales et des fibroblastes.