L'ARN interagissant avec Piwi ( piRNA ) est la plus grande classe de petites molécules d'ARN non codantes exprimées dans les cellules animales. Les piRNA forment des complexes ARN- protéine par le biais d'interactions avec les protéines Argonaute de la sous-famille piwi . Ces complexes piRNA sont principalement impliqués dans le silençage épigénétique et post-transcriptionnel des éléments transposables et d'autres transcrits parasites ou dérivés de répétitions, mais peuvent également être impliqués dans la régulation d'autres éléments génétiques dans les cellules germinales .
Les piRNA sont principalement créés à partir de loci qui fonctionnent comme des pièges à transposons qui fournissent une sorte d' immunité adaptative médiée par l'ARN contre les expansions et les invasions de transposons. Ils se distinguent des microARN (miRNA) par leur taille (26 à 31 nucléotides contre 21 à 24 nt), l'absence de conservation de séquence, une complexité accrue et l'indépendance de Dicer pour la biogenèse, au moins chez les animaux. ( La Dcl2 végétale peut jouer un rôle dans la biogenèse des rasi/piRNA.)
Les ARN double brin capables de faire taire les éléments répétés, alors connus sous le nom de petits ARN interférents associés aux répétitions (rasiRNA), ont été proposés chez la drosophile en 2001. En 2008, on ne savait toujours pas comment les piRNA étaient générés, mais des méthodes potentielles avaient été suggérées et il était certain que leur voie de biogenèse était distincte des miRNA et siRNA , tandis que le rasiRNA est désormais considéré comme une sous-espèce de piRNA.
Caractéristiques

Les piARN ont été identifiés chez les vertébrés et les invertébrés , et bien que la biogenèse et les modes d'action varient quelque peu entre les espèces, un certain nombre de caractéristiques sont conservées. Les piARN n'ont pas de motifs de structure secondaire clairs , en raison du fait que la longueur d'un piARN varie selon les espèces (de 21 à 31 nucléotides ), et le biais en faveur d'une uridine 5' est commun aux piARN chez les vertébrés et les invertébrés. Les piARN de Caenorhabditis elegans ont un monophosphate 5' et une modification 3' qui agit pour bloquer l'oxygène 2' ou 3' ; cela a également été confirmé chez Drosophila melanogaster , le poisson zèbre , la souris , et le rat . Cette modification 3' est une 2'-O-méthylation ; la raison de cette modification n'est pas claire, mais il a été suggéré qu'elle augmente la stabilité du piRNA.
Plus de 50 000 séquences uniques de piRNA ont été découvertes chez la souris et plus de 13 000 chez D. melanogaster . On pense qu'il existe plusieurs centaines de milliers d'espèces différentes de piRNA chez les mammifères .
Histoire et lieux
Au début des années 1980, on a découvert qu'une seule mutation dans le génome de la mouche à fruits pouvait activer spécifiquement toutes les copies d'un élément de type rétrovirus appelé Gypsy dans la lignée germinale femelle . Le site des mutations qui faisaient « danser » ces Gitans a ainsi été appelé le locus flamenco . En 2001, Aravin et al. ont proposé que le silençage médié par l'ARN double brin (ds) soit impliqué dans le contrôle des rétrotransposons dans la lignée germinale et en 2003, l'idée a émergé que les vestiges de transposons pourraient produire les ARNdb nécessaires au silençage des transposons « vivants ». Le séquençage du locus flamenco de 200 000 pb a été difficile, car il s'est avéré être rempli de fragments d'éléments transposables (104 insertions de 42 transposons différents, dont plusieurs Gitans), tous orientés dans la même direction. En effet, les piARN sont tous présents dans des groupes dans les génomes animaux ; ces groupes peuvent contenir aussi peu que dix ou plusieurs milliers de piARN correspondant à différents fragments de transposons phasés. Cela a conduit à l'idée en 2007 que dans les lignées germinales, un pool de piARN primaires est traité à partir de longs transcrits monocaténaires codés par des groupes de piARN dans l'orientation opposée des transposons, de sorte que les piARN peuvent s'hybrider et compléter les transcrits codés par les transposons, déclenchant ainsi leur dégradation. Tout transposon atterrissant dans la bonne orientation dans un tel groupe rendra l'individu plus ou moins immunisé contre ce transposon, et une telle mutation avantageuse se propagera rapidement dans la population. Les mutations originales du locus flamenco ont inhibé la transcription du transcrit maître, désactivant ainsi ce système de défense.
Un exemple historique d'invasion et de réponse Piwi est connu : le transposon de l'élément P a envahi un génome de Drosophila melanogaster au milieu du 20e siècle et, par croisement, en quelques décennies, toutes les mouches à fruits sauvages du monde (mais pas les souches de laboratoire isolées sur le plan reproductif) contenaient le même élément P. La répression d'une activité supplémentaire de l'élément P, se propageant presque simultanément, semble avoir eu lieu par la voie de l'ARN interagissant avec Piwi.
Les groupes de piARN dans les génomes peuvent désormais être facilement détectés via des méthodes bioinformatiques . Alors que les piARN de D. melanogaster et des vertébrés ont été localisés dans des zones dépourvues de gènes codant pour des protéines , des piARN de C. elegans ont été identifiés parmi des gènes codant pour des protéines.
Chez les mammifères, les piARN se trouvent à la fois dans les testicules et dans les ovaires , bien qu'ils ne semblent nécessaires que chez les mâles lignées germinales mâles et femelles .
Au niveau cellulaire, des piARN ont été trouvés à la fois dans le noyau et dans le cytoplasme , ce qui suggère que les voies piARN peuvent fonctionner dans ces deux zones et, par conséquent, peuvent avoir de multiples effets.
Classification
Il existe au moins trois sous-familles d'Argonautes (Ago) qui ont été trouvées chez les eucaryotes . Contrairement à la sous-famille Ago qui est présente chez les animaux, les plantes et la levure de fission, la sous-famille Piwi n'a été trouvée que chez les animaux. Le RasiRNA a été observé chez la drosophile et certains eucaryotes unicellulaires, mais sa présence chez les mammifères n'a pas été déterminée, contrairement au piRNA qui a été observé chez de nombreuses espèces d'invertébrés et de vertébrés, y compris les mammifères ; cependant, comme les protéines qui s'associent au rasiRNA se trouvent à la fois chez les vertébrés et les invertébrés, il est possible que des rasiRNA actifs existent et n'aient pas encore été observés chez d'autres animaux. Des RasiRNA ont été observés chez Schizosaccharomyces pombe , une espèce de levure, ainsi que dans certaines plantes, dont aucune n'a été observée comme contenant la sous-famille Piwi de protéines Argonaute. Il a été observé que le rasiRNA et le piRNA sont tous deux liés à la mère, mais plus spécifiquement, c'est la sous-famille de protéines Piwi qui est liée à la mère et conduit donc à l'observation que le rasiRNA et le piRNA sont liés à la mère.
Biogenèse
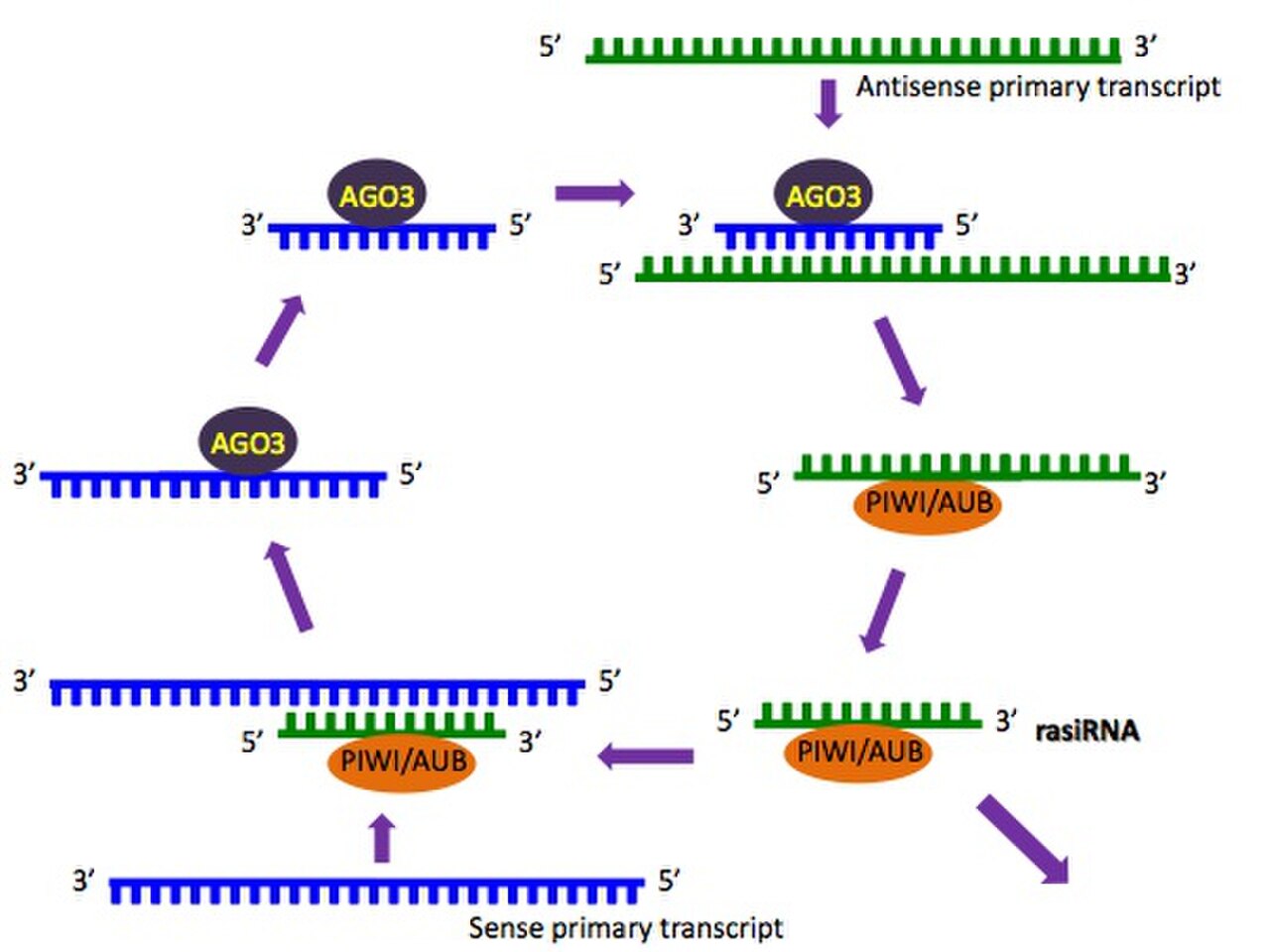
La biogenèse des piARN n'est pas encore entièrement comprise, bien que des mécanismes possibles aient été proposés. Les piARN présentent un biais de brin significatif, c'est-à-dire qu'ils sont dérivés d'un seul brin d' ADN , et cela peut indiquer qu'ils sont le produit de longues molécules précurseurs monocaténaires. Une voie de traitement primaire est suggérée comme étant la seule voie utilisée pour produire des piARN pachytènes ; dans ce mécanisme, les précurseurs de piARN sont transcrits, ce qui donne des piARN ayant tendance à cibler les uridines 5' . Un mécanisme de « ping-pong » est également proposé dans lequel les piARN primaires reconnaissent leurs cibles complémentaires et provoquent le recrutement de protéines piwi . Cela entraîne le clivage du transcrit à un point situé à dix nucléotides de l'extrémité 5' du piARN primaire, produisant le piARN secondaire. Ces piARN secondaires sont ciblés vers des séquences qui possèdent une adénine en dixième position. Étant donné que le piRNA impliqué dans le cycle ping-pong dirige ses attaques sur les transposons, le cycle ping-pong agit uniquement au niveau de la transcription . L'un ou les deux de ces mécanismes peuvent agir dans différentes espèces ; C. elegans , par exemple, possède des piRNA, mais ne semble pas du tout utiliser le mécanisme ping-pong.
Un nombre significatif de piRNAs identifiés chez le poisson zèbre et D. melanogaster contiennent de l'adénine en dixième position, et cela a été interprété comme une preuve possible d'un mécanisme de biosynthèse conservé à travers les espèces. Des signatures ping-pong ont été identifiées chez des animaux très primitifs tels que les éponges et les cnidaires, indiquant l'existence du cycle ping-pong déjà dans les premières branches des métazoaires.
Ping-pong
La voie piRNA Ping-Pong a été proposée pour la première fois à partir d'études sur la drosophile où le piRNA associé aux deux protéines cytoplasmiques Piwi, Aubergine (Aub) et Argonaute-3 (Ago3) présentait une fréquence élevée de complémentarité de séquence sur exactement 10 nucléotides à leurs extrémités 5'. Cette relation est connue sous le nom de « signature ping-pong » et est également observée dans le piRNA associé aux protéines Mili et Miwi2 isolées des testicules de souris. La fonction proposée de Ping-Pong chez la drosophile ou la souris reste à comprendre, mais une hypothèse principale est que l'interaction entre Aub et Ago3 permet un raffinement cyclique des piRNA qui sont les mieux adaptés pour cibler les séquences de transposons actifs. Les piRNA Aub sont principalement antisens aux transcrits d'éléments transposables et sont considérés comme le principal facteur de ciblage des transcrits délétères par complémentarité. A l'inverse, les séquences d'ARNpi d'Ago3 sont principalement orientées dans le sens des transcrits d'éléments transposables et sont dérivées du produit du clivage d'Aub de l'ARNm du transposon. Ainsi, l'ARNpi d'Ago3 n'a pas la capacité de cibler directement les transcrits d'éléments transposables. Par conséquent, il a été proposé que l'ARNpi d'Ago3 guide la production d'ARNpi chargés dans Aub en ciblant les transcrits de groupe d'ARNpi nouvellement exportés. Plusieurs éléments de preuve soutiennent l'effet d'Ago3 sur la production d'ARNpi d'Aub, en particulier en examinant le répertoire d'ARNpi dans les ovaires de drosophile qui sont mutants pour Ago3 et la protéine du domaine Tudor Kumo/Qin.
Le mécanisme moléculaire qui sous-tend le ping-pong implique probablement plusieurs facteurs associés à la voie piRNA. Il a été rapporté que Qin coordonnait le chargement d'Ago3 avec le piRNA, en plus d'interagir avec Aub et Ago3. Cependant, il a également été démontré que la protéine Tudor krimper ( A1ZAC4 ) interagissait avec Aub et Ago3 via ses domaines Tudor tout en se liant via son domaine N-terminal Krimper. Plus précisément, Krimper interagit avec Ago3 dans son état déchargé en piRNA, tandis que son interaction avec Aub dépend de la diméthylation symétrique des résidus d'arginine dans la région N-terminale d'Aub. Dans les cellules germinales Silkmoth, il a été proposé que la protéine Vasa coordonne le mécanisme Ping-Pong de Silkmoth Aub (Siwi) et Ago3.
Il est probable que le mécanisme du ping-pong soit principalement coordonné par Krimper, mais des facteurs tels que Kumo/Qin et Vasa, en plus d'autres facteurs, ont des fonctions nécessaires dans le mécanisme du ping-pong.
Mise en phase du piRNA
La voie piRNA de la drosophile peut être séparée en deux branches : la branche cytoplasmique composée d'Aub et d'Ago3 exploitant le mécanisme Ping-Pong, et la branche nucléaire, relative au silençage co-transcriptionnel des loci génomiques par Piwi dans le noyau. Grâce à des stratégies complémentaires, deux études montrent que le clivage de la cible par Aub et Ago3 déclenche le chargement « phasé » de piRNA dans Piwi. Le phasage commence par le ciblage et le clivage d'une cible complémentaire par Aub ou Ago3 associé à un piRNA « répondeur ». Une fois clivé, le transcrit ciblé est ensuite traité plus avant par un mécanisme qui nécessiterait l'endonucléase associée aux mitochondries, Zucchini, qui conduit au chargement de la protéine Piwi avec des fragments séquentiels du transcrit ciblé. De cette façon, la séquence piRNA « répondante » d'Aub ou d'Ago3 coupe une cible complémentaire qui est ensuite découpée à intervalles périodiques d'environ 27 nucléotides qui sont chargés séquentiellement dans la protéine Piwi. Une fois chargée de piRNA, Piwi pénètre ensuite dans le noyau de la cellule germinale pour faire taire de manière co-transcriptionnelle les transcrits naissants en complémentarité avec son guide piRNA. On ne sait pas actuellement si le phasage se produit dans d'autres organismes.
Fonction
La grande variation des séquences de piRNA et de la fonction de piwi selon les espèces contribue à la difficulté d'établir la fonctionnalité des piRNA. Cependant, comme d'autres petits ARN , on pense que les piRNA sont impliqués dans le silençage génique , en particulier le silençage des transposons . La majorité des piRNA sont antisens aux séquences de transposons, suggérant que les transposons sont les cibles des piRNA. Chez les mammifères, il semble que l'activité des piRNA dans le silençage des transposons soit la plus importante pendant le développement de l' embryon , et chez C. elegans et les humains, les piRNA sont nécessaires à la spermatogenèse .
Silençage de l'ARN
Le piARN joue un rôle dans le silençage de l'ARN via la formation d'un complexe de silençage induit par l'ARN (RISC). Les piARN interagissent avec les protéines piwi qui font partie d'une famille de protéines appelées les Argonautes . Celles-ci sont actives dans les testicules des mammifères et sont nécessaires au développement des cellules germinales et des cellules souches chez les invertébrés . Trois protéines de la sous-famille piwi - MIWI, MIWI2 et MILI - se sont avérées essentielles à la spermatogenèse chez la souris. Les piARN dirigent les protéines piwi vers leurs cibles de transposon. Une diminution ou une absence d' expression du gène PIWI est corrélée à une expression accrue des transposons. Les transposons ont un potentiel élevé pour provoquer des effets délétères sur leurs hôtes et, en fait, des mutations dans les voies piARN se sont avérées réduire la fertilité chez D. melanogaster . De plus, on pense que le piRNA et le petit ARN interférent endogène (endo-siRNA) peuvent avoir une fonctionnalité comparable et même redondante dans le contrôle des transposons dans les ovocytes de mammifères .
Les piARN semblent affecter des méthyltransférases particulières qui effectuent les méthylations nécessaires à la reconnaissance et au silence des transposons, mais cette relation n'est pas bien comprise.
Effets antiviraux
Chez les diptères, les piARN viraux dérivés de virus à ARN positif ont été identifiés pour la première fois dans les cellules somatiques ovariennes (OSS) de Drosophila . Des études expérimentales ultérieures ont démontré que la voie piARN n'est pas nécessaire à la défense antivirale chez Drosophila melanogaster . Cependant, chez les moustiques, la famille de protéines PIWI s'est élargie et certaines protéines PIWI ont été identifiées comme antivirales telles que Piwi4. Ainsi, les infections virales chez les moustiques produisent généralement des piARN dérivés de virus dans divers virus à ARN positif, à ARN négatif et à ADN simple brin.
Effets épigénétiques
Les piARN peuvent être transmis par voie maternelle, et, d'après les recherches menées sur D. melanogaster , les piARN peuvent être impliqués dans les effets épigénétiques d'origine maternelle . L'activité de piARN spécifiques dans le processus épigénétique nécessite également des interactions entre les protéines piwi et HP1a, ainsi que d'autres facteurs.
Protéines accessoires de la voie piRNA
Des tests génétiques examinant les défauts de fertilité ont permis d'identifier un certain nombre de protéines qui ne sont pas des Argonautes du clade Piwi, mais qui produisent les mêmes phénotypes de stérilité que les mutants Piwi.
DrosophileProtéines du domaine Tudor
De nombreux facteurs nécessaires à la voie piRNA chez Drosophila contiennent des domaines Tudor qui sont connus pour se lier aux résidus d'arginine diméthylés symétriquement (sDMA) présents dans les motifs de méthylation des protéines Piwi. Les protéines Piwi sont diméthylées symétriquement par le complexe méthylosome PRMT5, composé de Valois (MEP50) et de Capsulèen (dart5 ; PRMT5).
- Tudor (Tud)
- Qin/Kumo
- Broche E (SpnE)
- Crimpeur
- Tejas (Tej)
- Vreteno (Vret)
- Papi
- Yb ( fs(1)Yb )
- Frère de Yb (BoYB)
- Sœur de Yb (SoYB)
Non-TudorDrosophileProtéines de la voie piRNA
- Vasa
- Maelström (Mael)
Drosophileprotéines de la voie nucléaire des piRNA
- Rhinocéros (HP1D)
- Impasse
- Couper
- SetDB1 (Sans œuf)
- SuVar3–9
Enquête
Des avancées majeures dans l'étude des piARN ont été réalisées grâce à l'utilisation de techniques de séquençage de nouvelle génération , telles que le séquençage de la plateforme Solexa, 454 et Illumina . Ces techniques permettent d'analyser des populations d'ARN très complexes et hétérogènes comme les piARN. En raison de leur petite taille, l'expression et l'amplification des petits ARN peuvent être difficiles, c'est pourquoi des méthodes spécialisées basées sur la PCR ont été développées en réponse à cette difficulté. Cependant, la recherche a également révélé qu'un certain nombre de piARN annotés peuvent être des faux positifs ; par exemple, une majorité de piARN exprimés dans des tissus somatiques non gonadiques étaient considérés comme dérivés de fragments d'ARN non codants.