La proinsuline est le précurseur prohormonal de l'insuline produit dans les cellules bêta des îlots pancréatiques , des régions spécialisées du pancréas . Chez l'homme, la proinsuline est codée par le gène INS . Les îlots pancréatiques ne sécrètent qu'entre 1 % et 3 % de proinsuline intacte. Cependant, comme la proinsuline a une demi-vie plus longue que l'insuline, elle peut représenter entre 5 et 30 % des structures analogues à l'insuline circulant dans le sang. Les concentrations de proinsuline sont plus élevées après les repas et plus faibles lorsqu'une personne jeûne. De plus, bien que la proinsuline et l'insuline présentent des différences structurelles, la proinsuline présente une certaine affinité pour le récepteur de l'insuline . En raison des similitudes relatives de structure, la proinsuline peut produire entre 5 % et 10 % de l'activité métabolique induite de manière similaire par l'insuline.
La proinsuline est la dernière structure protéique à chaîne unique sécrétée par les cellules avant le clivage en insuline mature. La proinsuline a été découverte par le professeur Donald F. Steiner de l'Université de Chicago en 1967.
Structure
La proinsuline est constituée de 86 résidus chez l'homme (81 chez la vache), et formée de trois chaînes distinctes. La chaîne A, la chaîne B et la zone reliant les deux sont appelées peptide C. La structure correcte de la proinsuline est cruciale pour le repliement correct de l'insuline mature , car le placement du peptide C permet à la molécule de créer des liaisons disulfures correctement positionnées dans et entre les chaînes A et B. Il existe trois liaisons disulfures nécessaires pour que l'insuline mature ait la structure correcte. Deux de ces liaisons disulfures se trouvent entre les chaînes A et B, et une est une liaison intra-chaîne A. Les liaisons disulfures se produisent entre les septièmes résidus des chaînes A et B, le 20e résidu de la chaîne A et le 19e résidu de la chaîne B, et les 6e et 11e résidus de la chaîne A.
Le peptide C se situe entre les chaînes A et B de la proinsuline. La connexion entre la chaîne A et le peptide C est beaucoup plus stable que la jonction entre le peptide C et la chaîne B, avec des caractéristiques hélicoïdales alpha présentes à proximité de la connexion peptide C-chaîne A. La jonction peptide C-chaîne A se situe entre les résidus 64 et 65 de la proinsuline. Il s'agit respectivement de molécules de lysine et d'arginine . La connexion peptide C-chaîne B se situe entre deux résidus d'arginine aux positions 31 et 32 de la proinsuline.
Il existe une conservation d'une grande partie de la structure de la proinsuline parmi les espèces de mammifères, la plupart des changements de résidus observés d'une espèce à l'autre étant présents dans le peptide C. Cela dit, les résidus du peptide C qui sont conservés d'une espèce à l'autre interagissent avec des résidus conservés de manière similaire sur les chaînes A et B. Ainsi, on émet l'hypothèse que ces résidus conservés sont importants pour la fonctionnalité de l'insuline mature.
Synthèse et modification post-traductionnelle
La proinsuline est synthétisée sur des ribosomes associés à la membrane situés sur le réticulum endoplasmique rugueux , où elle est repliée et ses liaisons disulfures sont oxydées. Elle est ensuite transportée vers l' appareil de Golgi où elle est conditionnée dans des vésicules sécrétoires, et où elle est traitée par une série de protéases pour former l'insuline mature . L'insuline mature a 35 acides aminés de moins ; 4 sont complètement éliminés et les 31 restants forment le peptide C. Le peptide C est extrait du centre de la séquence de la proinsuline ; les deux autres extrémités (la chaîne B et la chaîne A) restent connectées par des liaisons disulfures.
La modification post-traductionnelle de la proinsuline en insuline mature ne se produit que dans les cellules bêta des îlots pancréatiques. Lorsque la proinsuline est transportée à travers l'appareil de Golgi, le peptide C est clivé. Ce clivage se produit à l'aide de deux endoprotéases. Les endoprotéases de type I , PC1 et PC3, perturbent la connexion peptide C-chaîne B. PC2, une endoprotéase de type II, coupe la liaison peptide C-chaîne A. La molécule résultante, maintenant l'insuline mature, est stockée sous forme d'hexamère dans des vésicules sécrétoires et est stabilisée avec des ions jusqu'à ce qu'elle soit sécrétée.
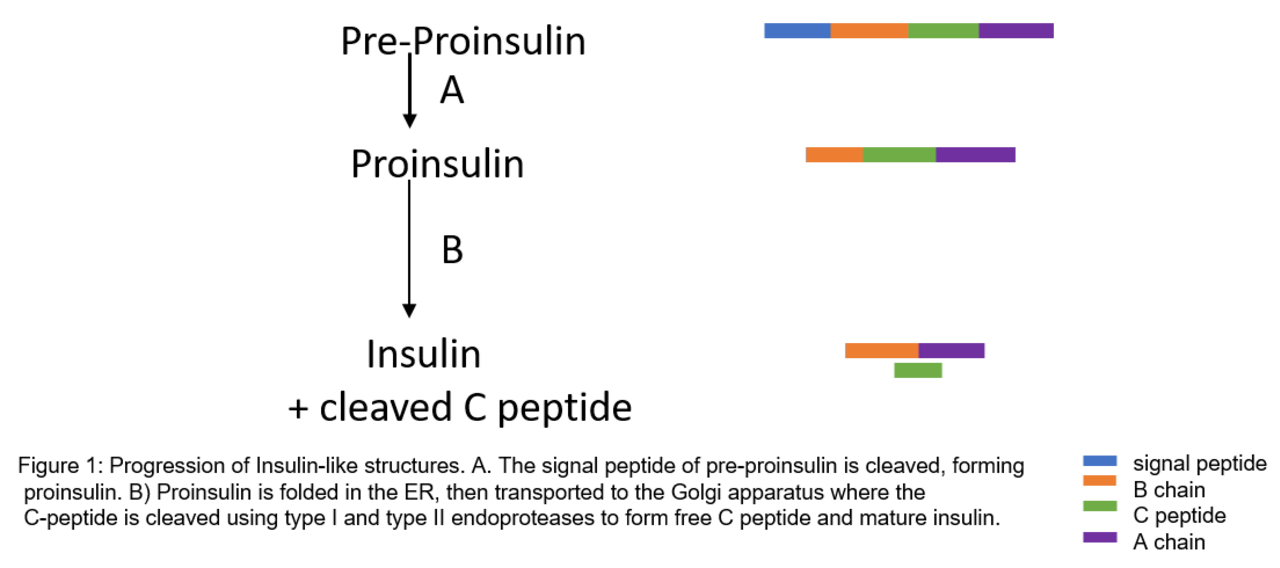
Immunogénicité
Lorsque l'insuline a été initialement purifiée à partir de pancréas bovin ou porcin , toute la proinsuline n'a pas été complètement éliminée. Lorsque certaines personnes ont utilisé ces insulines, la proinsuline a pu provoquer une réaction du corps avec une éruption cutanée, une résistance à l'insuline ou même la formation de bosses ou de bosses sur la peau à l'endroit où l'insuline a été injectée. Cela peut être décrit comme une lésion iatrogène due à de légères différences entre la proinsuline des différentes espèces. Depuis la fin des années 1970, lorsque l'insuline porcine hautement purifiée a été introduite et que le niveau de pureté de l'insuline a atteint 99 %, cela a cessé d'être un problème clinique important. En ce qui concerne leur influence sur la pharmacocinétique de l'insuline, des concentrations modérées de certains anticorps anti-insuline peuvent être bénéfiques pour tous les diabétiques sans sécrétion endogène d'insuline (par exemple les personnes atteintes de diabète de type 1 ) car les anticorps de liaison à l'insuline augmentent efficacement le taux de clairance et l'espace de distribution de l'insuline et aident à prolonger ses demi-vies pharmacologiques et biologiques.
Pertinence médicale
Historiquement, de nombreuses maladies métaboliques liées à l'insuline se sont concentrées sur l'insuline mature. Cependant, ces dernières années, l'importance d'étudier la structure et la fonction de la proinsuline ou du rapport proinsuline:insuline en relation avec ces maladies est devenue de plus en plus évidente.
Des niveaux accrus de proinsuline dans le système circulatoire par rapport aux concentrations d'insuline mature peuvent indiquer une résistance imminente à l'insuline et le développement d' un diabète de type 2. D'autres problèmes liés à la proinsuline pouvant conduire au diabète comprennent des mutations dans le nombre de cystéines présentes, qui pourraient affecter le repliement correct. Si la mutation ne provoque qu'un changement léger, elle pourrait simplement stresser la capacité du réticulum endoplasmique à replier correctement la protéine. Ce stress, après un certain temps, entraînerait une diminution du nombre de cellules bêta produisant de l'insuline mature, et conduirait alors au diabète sucré.
La proinsuline postnatale est essentielle à la régulation métabolique. Cependant, la proinsuline chez les nouveau-nés est importante pour le développement normal des nerfs oculaires, le développement du cœur et la survie générale des cellules embryonnaires. La régulation de la concentration de proinsuline pendant le développement embryonnaire est cruciale, car une quantité trop importante ou trop faible de peptide peut provoquer des malformations et la mort du fœtus. Jusqu'à présent, dans l'étude du diabète sucré néonatal, seules les mutations de changement d'acides aminés trouvées dans le domaine B conduisent à la maladie.