La pyroptose est une forme hautement inflammatoire de mort cellulaire lytique programmée qui survient le plus souvent lors d'une infection par des agents pathogènes intracellulaires et qui est susceptible de faire partie de la réponse antimicrobienne . Ce processus favorise l'élimination rapide de diverses infections bactériennes, virales, fongiques et protozoaires en supprimant les niches de réplication intracellulaire et en renforçant les réponses défensives de l'hôte. La pyroptose peut se produire dans les cellules immunitaires et se produit également dans les kératinocytes et certaines cellules épithéliales .
Le processus est initié par la formation d'un grand complexe supramoléculaire appelé inflammasome (également connu sous le nom de pyroptosome) lors de signaux de danger intracellulaires. L'inflammasome active un ensemble différent de caspases par rapport à l'apoptose , par exemple, la caspase-1/4/5 chez l'homme et la caspase-11 chez la souris. Ces caspases contribuent à la maturation et à l'activation des cytokines pro-inflammatoires IL-1β et IL-18, ainsi que de la protéine gasdermine D formant des pores . La formation de pores provoque la rupture de la membrane cellulaire et la libération de cytokines, ainsi que de diverses molécules de motifs moléculaires associés aux dommages (DAMP) telles que HMGB-1, ATP et ADN, hors de la cellule. Ces molécules recrutent davantage de cellules immunitaires et perpétuent davantage la cascade inflammatoire dans le tissu.
Cependant, dans les maladies chroniques pathogènes, la réponse inflammatoire n'éradique pas le stimulus primaire. Une forme chronique d'inflammation s'ensuit qui contribue finalement aux lésions tissulaires. La pyroptose est associée à des maladies telles que les maladies auto-inflammatoires, métaboliques et cardiovasculaires , ainsi qu'au cancer et à la neurodégénérescence . Parmi les exemples de pyroptose, on peut citer la mort cellulaire induite dans les macrophages infectés par Salmonella et les cellules T auxiliaires infectées par le VIH qui ont avorté .
Découverte
Ce type de mort cellulaire programmée intrinsèquement pro-inflammatoire a été nommé pyroptose en 2001 par Molly Brennan et le Dr Brad T. Cookson, professeur associé de microbiologie et de médecine de laboratoire à l' Université de Washington . Le grec pyro fait référence au feu et ptosis signifie chute. Le terme composé de pyroptose peut être compris comme « chute ardente », qui décrit l'éclatement des signaux chimiques pro-inflammatoires de la cellule mourante. La pyroptose a une morphologie et un mécanisme distincts par rapport à ceux d'autres formes de mort cellulaire. Il a été suggéré que l'infection microbienne était la principale pression évolutive pour cette voie. La formation d'inflammasome était initialement considérée comme nécessaire à l'induction de la pyroptose, mais en 2013, la voie non canonique dépendante de la caspase-11 a été découverte, suggérant que les lipopolysaccharides (LPS) peuvent déclencher la pyroptose et les réponses inflammatoires ultérieures indépendamment du récepteur de type Toll 4 (TLR4) . En 2015, la gasdermine D (GSDMD) a été identifiée comme l'effecteur de la pyroptose qui forme des pores dans la membrane cellulaire. En 2021, la structure haute résolution du pore GSDMD a été résolue par cryomicroscopie électronique (cryo-EM). Également en 2021, une molécule supplémentaire, NINJ1 , s'est avérée nécessaire à la rupture de la membrane plasmique lors de la pyroptose.
Caractéristiques morphologiques
La pyroptose, en tant que forme de mort cellulaire programmée, présente de nombreuses différences morphologiques par rapport à l'apoptose . La pyroptose et l'apoptose subissent toutes deux une condensation de la chromatine , mais pendant l'apoptose, le noyau se décompose en plusieurs corps de chromatine ; dans le cas de la pyroptose, le noyau reste intact. Dans une cellule qui subit une pyroptose, des pores de gasdermine se forment sur la membrane plasmique, ce qui entraîne un afflux d'eau.
En termes de mécanisme, la pyroptose est activée par des caspases inflammatoires, notamment la caspase-1/4/5 chez l'homme et la caspase-11 chez la souris. La caspase-8 peut agir comme un régulateur en amont de l'activation de l'inflammasome de manière dépendante du contexte. de la caspase-3 peut avoir lieu à la fois lors de l'apoptose et de la pyroptose.
Bien que la pyroptose et la nécroptose soient toutes deux déclenchées par la formation de pores membranaires, la pyroptose est plus contrôlée. Les cellules qui subissent une pyroptose présentent des cloques membranaires et produisent des protubérances appelées corps pyroptotiques, un processus qui n'est pas observé dans la nécroptose. De plus, la nécroptose fonctionne de manière indépendante de la caspase. Il est proposé que la pyroptose et la nécroptose puissent agir comme des systèmes de défense contre les agents pathogènes lorsque les voies apoptotiques sont bloquées.
Mécanisme
Le système immunitaire inné , en utilisant des récepteurs de reconnaissance de motifs codés par la lignée germinale (PRR), peut reconnaître une large gamme de motifs moléculaires associés aux pathogènes (PAMP) et de motifs moléculaires associés aux dommages (DAMP) lors d'une infection microbienne. Les exemples classiques de PRR comprennent les récepteurs de type Toll (TLR) et les récepteurs de type NOD (NLR). inflammasomes complexes multiprotéiques , qui activent ensuite les caspases pour initier la pyroptose. La voie de l'inflammasome peut être canonique ou non canonique, la première utilisant des inflammasomes activant la caspase-1 et la seconde utilisant d'autres caspases.
La voie canonique de l'inflammasome
Dans la voie canonique de l'inflammasome, les PAMP et les DAMP sont reconnus par certains PRR endogènes. Par exemple, les protéines NLR NLRC4 peuvent reconnaître la flagelline et les composants du système de sécrétion de type III . NLRP3 est activée par des événements cellulaires induits par différents stimuli PAMP et DAMP. Certaines protéines non NLR comme l'AIM2 ( absent in melanoma 2 ) et la pyrine peuvent également être activées et former des inflammasomes. De plus, les PRR non formateurs d'inflammasomes tels que les TLR, NOD1 et NOD2 jouent également un rôle important dans la pyroptose. Ces récepteurs régulent à la hausse l'expression de cytokines inflammatoires telles que l'IFN α/β, le facteur de nécrose tumorale (TNF) , l'IL-6 et l' IL-12 via les voies de signalisation NF-κB et MAPK. De plus, la pro-IL-1β et la pro-IL-18 sont libérées pour être traitées par la caspase-1 médiée par la cystéine.
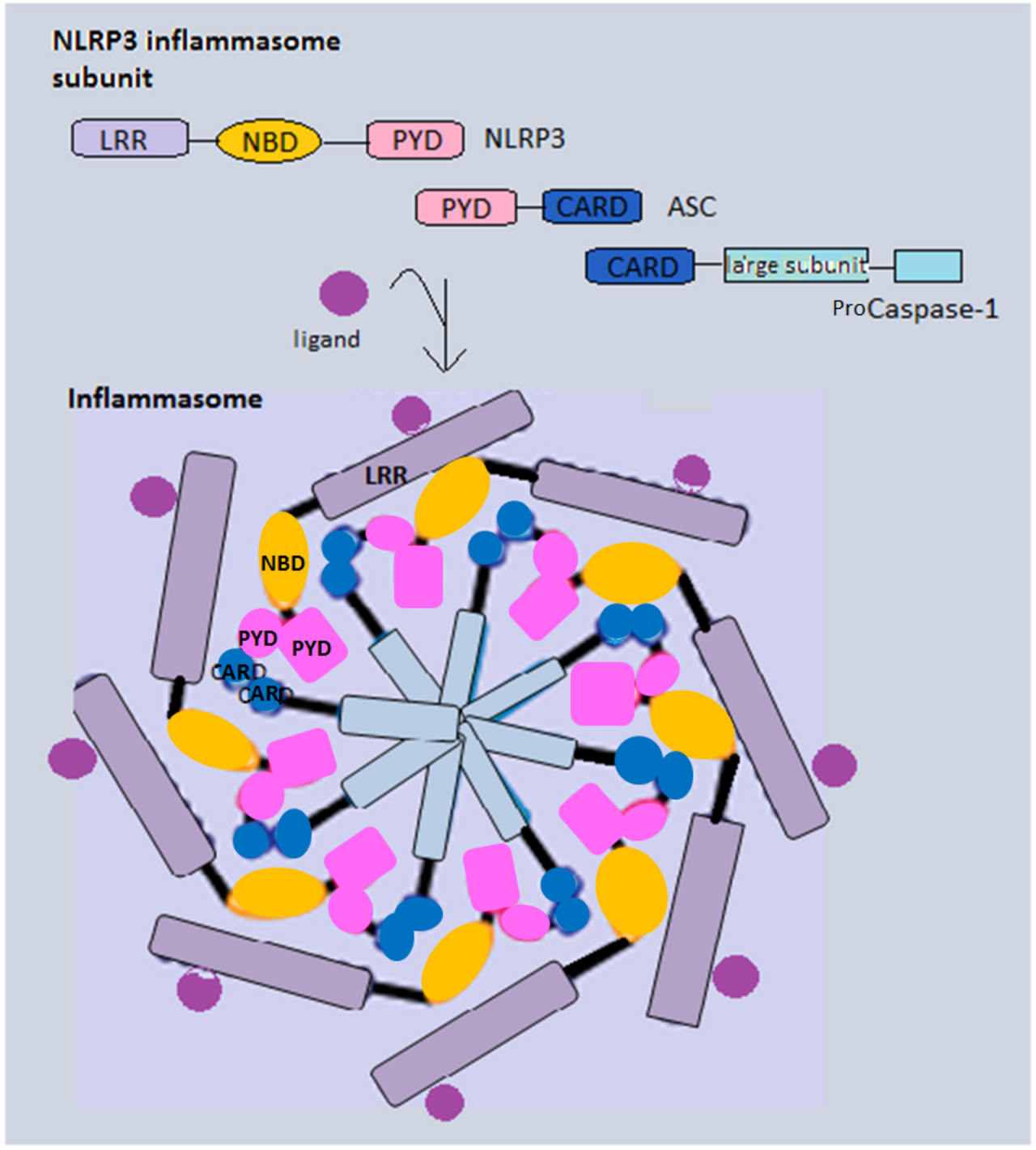
Français Les inflammasomes canoniques contiennent principalement trois composants : une protéine de détection (PRR), un adaptateur ( ASC ) et un effecteur ( caspase-1 ). En général, les protéines NLR formant des inflammasomes partagent une structure similaire, plusieurs domaines de répétition riche en leucine (LRR), un domaine central de liaison et d'oligomérisation des nucléotides (NBD) et un domaine pyrine N-terminal (PYD). NLRP3, par exemple, recrute la protéine adaptatrice ASC via l'interaction PYD-PYD. La pro-caspase-1 et l'ASC contiennent toutes deux un domaine d'activation et de recrutement de la caspase (CARD), et cette interaction homotypique CARD-CARD permet le clivage autocatalytique et le réassemblage de la procaspase-1 pour former la caspase-1 active. Alternativement, NLRC4 peut recruter directement la pro-caspase-1, car elle possède un CARD au lieu d'un PYD. En plus de leur formation en tant que complexe pour induire la pyroptose, les inflammasomes peuvent également être des composants à part entière de complexes plus grands induisant la mort cellulaire appelés PANoptosomes pour induire la PANoptose , une autre forme inflammatoire de mort cellulaire.
La caspase-1 activée est responsable du clivage de la pro-IL-1β et de la pro-IL-18. Ces cytokines, une fois traitées, seront sous leur forme biologiquement active, prêtes à être libérées des cellules hôtes. De plus, la caspase-1 clive également la gasdermine D cytosolique (GSDMD) . La GSDMD peut être clivée pour produire un domaine N-terminal (GSDMD-N) et un domaine C-terminal (GSDMD-C). La GSDMD-N peut s'oligomériser et former des pores transmembranaires d'un diamètre intérieur de 10 à 14 nm. Les pores permettent la sécrétion d'IL-1β et d'IL-18 et de divers contenus cytosoliques dans l'espace extracellulaire, et ils perturbent également le gradient ionique cellulaire. L'augmentation de la pression osmotique qui en résulte provoque un afflux d'eau suivi d'un gonflement et d'un éclatement des cellules. Notamment, GSDMD-N est auto-inhibé par le domaine C-terminal de GSDMD avant le clivage pour empêcher la lyse cellulaire dans des conditions normales. De plus, GSDMD-N ne peut s'insérer dans la membrane interne qu'avec des compositions lipidiques spécifiques, ce qui limite ses dommages aux cellules voisines. En aval de GSDMD, NINJ1 est maintenant considéré comme nécessaire à la rupture de la membrane plasmique pendant la pyroptose.
La voie non canonique de l'inflammasome
La voie de l'inflammasome non canonique est initiée par la liaison du lipopolysaccharide (LPS) des bactéries Gram-négatives directement sur la caspase-4/5 chez l'homme et la caspase-11 chez la souris . La liaison du LPS sur ces caspases favorise leur oligomérisation et leur activation. Ces caspases peuvent cliver la GSDMD pour libérer la GSDMD-N et déclencher la pyroptose. De plus, un afflux d'ions potassium lors de la perméabilisation de la membrane déclenche l'activation de NLRP3, qui conduit ensuite à la formation de l'inflammasome NLRP3 et à l'activation de la caspase-1. Ces processus facilitent le clivage de la GSDMD et favorisent la maturation et la libération de cytokines pro-inflammatoires.

Voie de mort cellulaire dépendante de la caspase-3
Une voie alternative reliant l'apoptose et la pyroptose a été récemment proposée. La caspase-3, une caspase exécutrice de l'apoptose, peut cliver la gasdermine E (GSDME) pour produire un fragment N-terminal et un fragment C-terminal d'une manière similaire au clivage de la GSDMD. Lorsque les cellules apoptotiques ne sont pas éliminées par les macrophages, l'expression de la GSDME est alors régulée à la hausse par p53. La GSDME est ensuite activée par la caspase-3 pour former des pores sur la membrane cellulaire. Il a également été découvert que la GSDME peut perméabiliser les membranes mitochondriales pour libérer le cytochrome c , qui active davantage la caspase-3 et accélère le clivage de la GSDME. Cette boucle de rétroaction positive garantit que la mort cellulaire programmée se poursuit.
Pertinence clinique
Infection
La pyroptose agit comme un mécanisme de défense contre l'infection en induisant une inflammation pathologique. La formation d'inflammasomes et l'activité de la caspase-1 déterminent l'équilibre entre la résolution du pathogène et la maladie.
Dans une cellule saine, l'activation de la caspase-1 permet de lutter contre l'infection causée par Salmonella et Shigella en introduisant la mort cellulaire pour limiter la croissance du pathogène. Lorsque le signal de « danger » est détecté, les cellules quiescentes seront activées pour subir une pyroptose et produire des cytokines inflammatoires IL-1β et IL-18. L'IL-18 stimulera la production d'IFNγ et initiera le développement des réponses T H 1. (Les réponses T H 1 ont tendance à libérer des cytokines qui dirigent une élimination immédiate du pathogène.) L'activation cellulaire entraîne une augmentation des niveaux de cytokines, ce qui augmentera les conséquences de l'inflammation et cela, à son tour, contribue au développement de la réponse adaptative à mesure que l'infection progresse. La résolution ultime éliminera les pathogènes.
En revanche, une inflammation persistante produira un excès de cellules immunitaires, ce qui est préjudiciable. Si les cycles d'amplification persistent, des troubles métaboliques, des maladies auto-inflammatoires et des lésions hépatiques associées à une inflammation chronique surviendront.
Récemment, la pyroptose et les voies en aval ont été identifiées comme des cibles prometteuses pour le traitement des maladies graves associées au COVID-19.
Maladie cérébrovasculaire
Des études récentes montrent que la pyroptose joue un rôle dans la physiopathologie de l’hémorragie intracérébrale, et l’atténuation de la pyroptose pourrait être une stratégie d’intervention pour inhiber la réponse inflammatoire après une hémorragie intracérébrale.
Cancer
La pyroptose, en tant que mort cellulaire programmée associée à l'inflammation, a de vastes implications dans divers types de cancer. Principalement, la pyroptose peut tuer les cellules cancéreuses et inhiber le développement tumoral en présence de DAMP endogènes. Dans certains cas, la GSDMD peut être utilisée comme marqueur pronostique des cancers. Cependant, la production prolongée de corps inflammatoires peut faciliter la formation de microenvironnements qui favorisent la croissance tumorale. La compréhension des mécanismes de la pyroptose et l'identification des molécules associées à la pyroptose peuvent être utiles dans le traitement de différents cancers.
Dans les cellules cancéreuses gastriques , la présence de GSDMD peut inhiber les complexes cycline A2 / CDK2 , ce qui entraîne un arrêt du cycle cellulaire et donc un développement tumoral inhibé. De plus, la concentration cellulaire de GSDME augmente lorsque les cellules cancéreuses gastriques sont traitées avec certains médicaments de chimiothérapie. Le GSDME active alors la caspase-3 et déclenche la mort cellulaire pyroptotique.
Le cancer du col de l'utérus peut être causé par une infection par le virus du papillome humain (HPV). La protéine AIM2 peut reconnaître l'ADN viral dans le cytoplasme et former l'inflammasome AIM2, qui se déclenche ensuite par une voie de pyroptose canonique dépendante de la caspase-1. L'infection par le HPV provoque la régulation positive de la protéine sirtuine 1 , qui perturbe le facteur de transcription pour AIM2, RelB . L'inactivation de la sirtuine 1 régule positivement l'expression d'AIM2 et déclenche la pyroptose.
Trouble métabolique
Le niveau d'expression de l'inflammasome NLRP3 et de la caspase-1 est directement lié à la gravité de plusieurs syndromes métaboliques, tels que l'obésité et le diabète sucré de type II (DT2) . En effet, le niveau de production ultérieur d'IL-1β et d'IL-18, des cytokines qui altèrent la sécrétion d' insuline , est affecté par l'activité de la caspase-1. Le niveau d'absorption du glucose est alors diminué, et la condition est connue sous le nom de résistance à l'insuline . La condition est encore accélérée par la destruction des cellules bêta pancréatiques induite par l'IL-1β .
Cryopyrinopathies
Une mutation dans le gène codant les inflammasomes conduit à un groupe de maladies auto-inflammatoires appelées cryopyrinopathies. Ce groupe comprend le syndrome de Muckle-Wells , le syndrome auto-inflammatoire du froid et le syndrome cutané et articulaire neurologique infantile chronique , tous présentant des symptômes de fièvres soudaines et d'inflammation localisée. Le gène muté dans ces cas est le NLRP3, qui empêche l'activation de l'inflammasome et entraîne une production excessive d'IL-1β. Cet effet est connu sous le nom de « gain de fonction ».
VIH et SIDA
Des études récentes démontrent que la pyroptose médiée par la caspase-1 entraîne une déplétion des lymphocytes T CD4 et une inflammation par le VIH , deux événements caractéristiques qui propulsent la progression de la maladie du VIH vers le SIDA . Bien que la pyroptose contribue à la capacité de l'hôte à limiter et à éliminer rapidement l'infection en supprimant les niches de réplication intracellulaire et en améliorant les réponses défensives par la libération de cytokines pro-inflammatoires et de signaux de danger endogènes, dans l'inflammation pathogène, telle que celle provoquée par le VIH-1, cette réponse bénéfique n'éradique pas le stimulus primaire. En fait, elle semble créer un cercle vicieux pathogène dans lequel les lymphocytes T CD4 mourants libèrent des signaux inflammatoires qui attirent davantage de cellules dans les tissus lymphoïdes infectés pour mourir et produire une inflammation chronique et des lésions tissulaires. Il pourrait être possible de briser ce cycle pathogène avec des inhibiteurs de la caspase-1 sûrs et efficaces. Ces agents pourraient former une nouvelle et passionnante thérapie « anti-SIDA » pour les sujets infectés par le VIH dans laquelle le traitement cible l'hôte plutôt que le virus. Il est à noter que les souris déficientes en caspase-1 se développent normalement, ce qui suggère que l'inhibition de cette protéine produirait des effets thérapeutiques bénéfiques plutôt que nocifs chez les patients atteints du VIH.