Un facteur sigma ( facteur σ ou facteur de spécificité ) est une protéine nécessaire à l'initiation de la transcription chez les bactéries . Il s'agit d'un facteur d'initiation de la transcription bactérienne qui permet la liaison spécifique de l'ARN polymérase (RNAP) aux promoteurs de gènes . Il est homologue au facteur de transcription B des archées et au facteur eucaryote TFIIB . Le facteur sigma spécifique utilisé pour initier la transcription d'un gène donné varie en fonction du gène et des signaux environnementaux nécessaires pour initier la transcription de ce gène. La sélection des promoteurs par l'ARN polymérase dépend du facteur sigma qui lui est associé. On les trouve également dans les chloroplastes des plantes en tant que partie de la polymérase codée par les plastes (PEP) de type bactérien.
Le facteur sigma, avec l'ARN polymérase, est connu sous le nom d'holoenzyme ARN polymérase . Chaque molécule d'holoenzyme ARN polymérase contient exactement une sous-unité de facteur sigma, qui dans la bactérie modèle Escherichia coli est l'une de celles énumérées ci-dessous. Le nombre de facteurs sigma varie selon les espèces bactériennes. E. coli possède sept facteurs sigma. Les facteurs sigma se distinguent par leurs poids moléculaires caractéristiques . Par exemple, σ 70 est le facteur sigma avec un poids moléculaire de 70 kDa .
Le facteur sigma dans le complexe holoenzyme de l'ARN polymérase est nécessaire à l'initiation de la transcription, bien qu'une fois cette étape terminée, il soit dissocié du complexe et l'ARN polymérase continue son élongation d'elle-même.
Facteurs sigma spécialisés
Différents facteurs sigma sont utilisés dans différentes conditions environnementales. Ces facteurs sigma spécialisés se lient aux promoteurs des gènes adaptés aux conditions environnementales, augmentant ainsi la transcription de ces gènes.
Facteurs sigma dans E. coli :
- σ70(RpoD) – σ A – le facteur sigma « de ménage » ou également appelé facteur sigma primaire (Groupe 1), transcrit la plupart des gènes dans les cellules en croissance. Chaque cellule possède un facteur sigma « de ménage » qui maintient les gènes et les voies essentiels en fonctionnement. Dans le cas d' E. coli et d'autres bactéries en forme de bâtonnet à Gram négatif, le facteur sigma « de ménage » est σ 70 . Les gènes reconnus par σ 70 contiennent tous des séquences consensus promotrices similaires constituées de deux parties. Par rapport à la base d'ADN correspondant au début de la transcription d'ARN, les séquences promotrices consensus sont généralement centrées à 10 et 35 nucléotides avant le début de la transcription (−10 et −35).
- σ19 (FecI) – le facteur sigma du citrate ferrique, régule le gène fec pour le transport et le métabolisme du fer
- σ 24 (RpoE) – réponse au stress thermique extrême et facteur sigma des protéines extracellulaires
- σ 28 (RpoF/FliA) – le facteur sigma de synthèse flagellaire et de chimiotaxie
- σ 32 (RpoH) – le facteur sigma de choc thermique , il est activé lorsque les bactéries sont exposées à la chaleur. En raison de son expression plus élevée, le facteur se lie avec une forte probabilité à l'enzyme polymérase-core. Ce faisant, d'autres protéines de choc thermique sont exprimées, ce qui permet à la cellule de survivre à des températures plus élevées. Certaines des enzymes exprimées lors de l'activation de σ 32 sont des chaperons , des protéases et des enzymes de réparation de l'ADN.
- σ 38 (RpoS) – le facteur sigma de la phase de famine/stationnaire
- σ 54 (RpoN) – le facteur sigma de limitation de l'azote
Il existe également des facteurs anti-sigma qui inhibent la fonction des facteurs sigma et des facteurs anti-anti-sigma qui restaurent la fonction du facteur sigma.
Structure
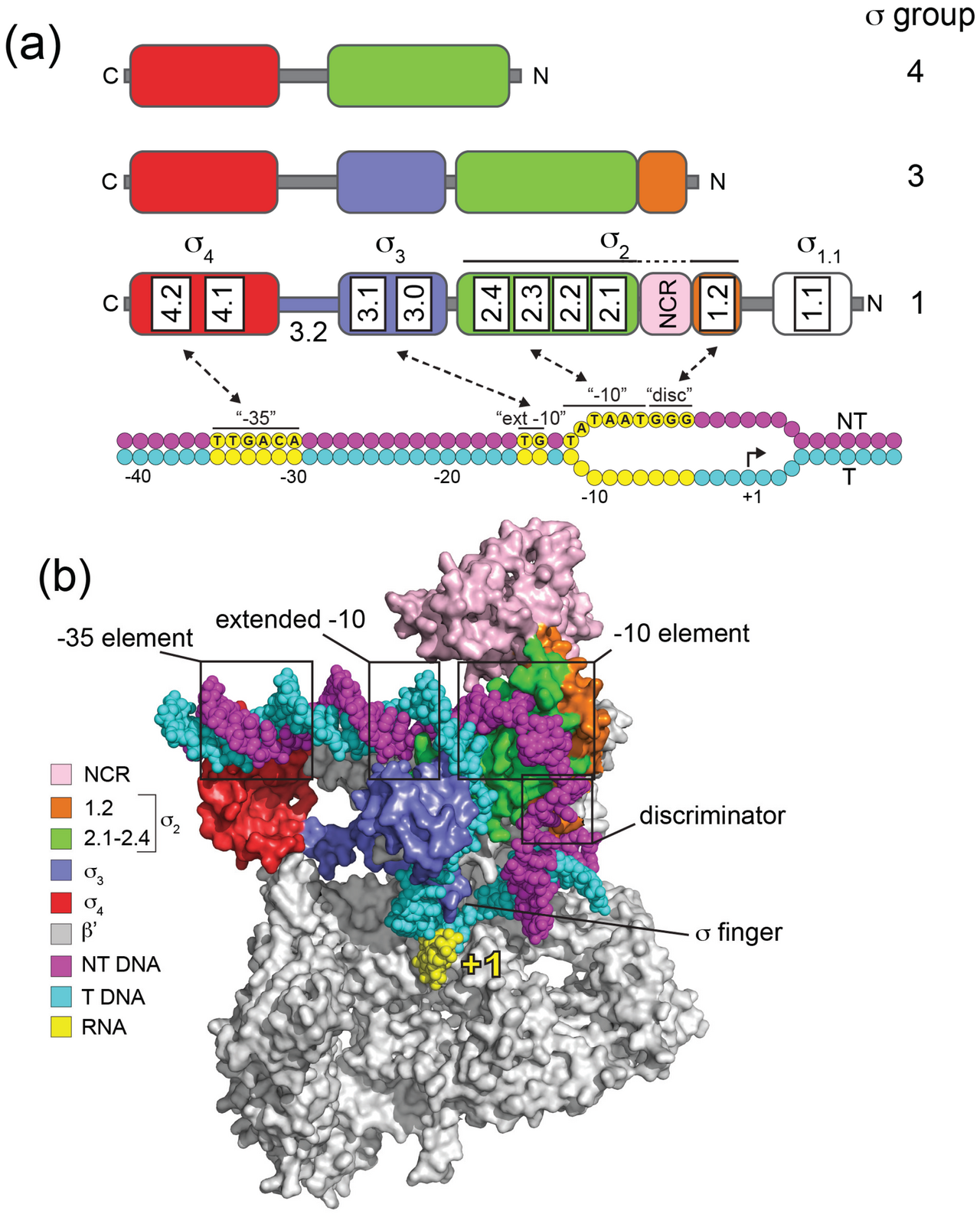
En raison de la similarité des séquences, la plupart des facteurs sigma sont de type σ 70 ( InterPro : IPR000943 ). Ils possèdent quatre régions principales (domaines) qui sont généralement conservées :
N-terminus --------------------- C-terminus 1.1 2 3 4
Les régions sont elles-mêmes subdivisées. Par exemple, la région 2 comprend les régions 1.2 et 2.1 à 2.4.
Le domaine 1.1 se trouve uniquement dans les « facteurs sigma primaires » (RpoD, RpoS dans E.coli ; « Groupe 1 »). Il permet de garantir que le facteur sigma ne se lie au promoteur que lorsqu'il est complexé avec l'ARN polymérase. Les domaines 2 à 4 interagissent chacun avec des éléments promoteurs spécifiques et avec l'ARN polymérase. La région 2.4 reconnaît et se lie à l'élément promoteur −10 (appelé « boîte Pribnow »). La région 4.2 reconnaît et se lie à l'élément promoteur −35.
Tous les facteurs sigma de la famille σ 70 ne contiennent pas tous les domaines. Le groupe 2, qui comprend RpoS, est très similaire au groupe 1 mais n'a pas le domaine 1. Le groupe 3 n'a pas non plus le domaine 1 et comprend σ 28. Le groupe 4, également connu sous le nom de groupe de fonction extracytoplasmique (ECF), n'a pas les domaines σ1.1 et σ3. RpoE en est un membre.
D'autres facteurs sigma connus sont de type σ 54 /RpoN ( InterPro : IPR000394 ). Ce sont des facteurs sigma fonctionnels, mais ils ont des séquences d'acides aminés primaires significativement différentes.
Rétention pendant l'élongation de la transcription
L'ARN polymérase centrale (composée de 2 sous-unités alpha (α), 1 sous-unité bêta (β), 1 sous-unité bêta-prime (β') et 1 sous-unité oméga (ω)) se lie à un facteur sigma pour former un complexe appelé holoenzyme ARN polymérase . On pensait auparavant que l'holoenzyme ARN polymérase initiait la transcription, tandis que l'ARN polymérase centrale synthétisait seule l'ARN. Ainsi, l'opinion acceptée était que le facteur sigma devait se dissocier lors de la transition de l'initiation de la transcription à l'élongation de la transcription (cette transition est appelée « échappement du promoteur »). Cette opinion était basée sur l'analyse de complexes purifiés d'ARN polymérase bloqués à l'initiation et à l'élongation. Enfin, les modèles structurels des complexes d'ARN polymérase prédisaient que, lorsque le produit d'ARN en croissance devenait plus long que ~15 nucléotides, le sigma devait être « poussé hors » de l'holoenzyme, car il y avait un conflit stérique entre l'ARN et un domaine sigma. Cependant, σ 70 peut rester lié en complexe avec l'ARN polymérase du noyau au début de l'élongation et parfois tout au long de l'élongation. En effet, le phénomène de pause proximale du promoteur indique que sigma joue un rôle au cours de l'élongation précoce. Toutes les études sont cohérentes avec l'hypothèse selon laquelle l'échappement du promoteur réduit la durée de vie de l'interaction sigma-noyau de très longue au début (trop longue pour être mesurée dans une expérience biochimique typique) à une durée de vie plus courte et mesurable lors de la transition vers l'élongation.
Cycle Sigma
On a longtemps pensé que le facteur sigma quittait obligatoirement l'enzyme centrale une fois qu'il avait initié la transcription, lui permettant de se lier à une autre enzyme centrale et d'initier la transcription à un autre site. Ainsi, le facteur sigma passerait d'un noyau à un autre. Cependant, le transfert d'énergie par résonance de fluorescence a été utilisé pour montrer que le facteur sigma ne quitte pas obligatoirement le noyau. Au lieu de cela, il change sa liaison avec le noyau pendant l'initiation et l'élongation. Par conséquent, le facteur sigma alterne entre un état fortement lié pendant l'initiation et un état faiblement lié pendant l'élongation.
Concours de facteur Sigma
Il a été démontré que le nombre de RNAP dans les cellules bactériennes (par exemple, E. coli ) est inférieur au nombre de facteurs sigma. Par conséquent, si un certain facteur sigma est surexprimé, non seulement cela augmentera les niveaux d'expression des gènes dont les promoteurs ont une préférence pour ce facteur sigma, mais cela réduira également la probabilité que des gènes dont les promoteurs ont une préférence pour d'autres facteurs sigma soient exprimés.
L'initiation de la transcription comporte deux étapes principales limitant la vitesse : la formation du complexe fermé et la formation du complexe ouvert. Cependant, seule la dynamique de la première étape dépend de la concentration des facteurs sigma. Il est intéressant de noter que plus la formation du complexe fermé est rapide par rapport à la formation du complexe ouvert, moins un promoteur est sensible aux changements de concentration des facteurs sigma (voir pour un modèle et des données empiriques sur ce phénomène).
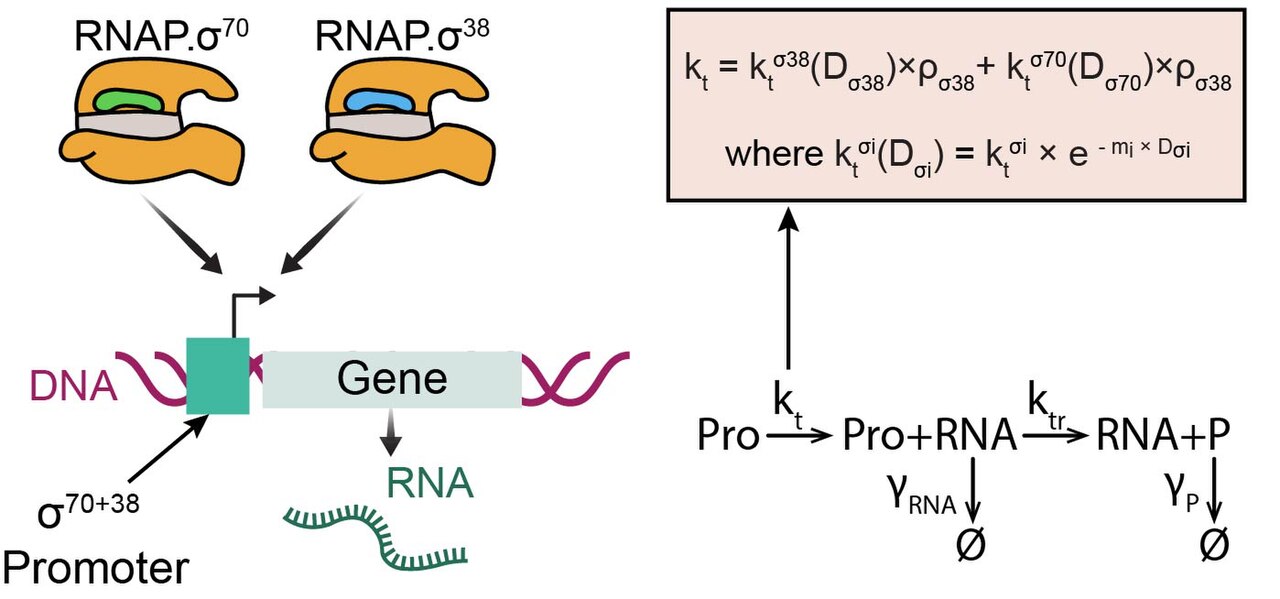
Gènes avec une préférence pour le double facteur sigma
Alors que la plupart des gènes d' E. coli peuvent être reconnus par une RNAP avec un et un seul type de facteur sigma (par exemple sigma 70), quelques gènes (~ 5 %) ont ce qu'on appelle une « préférence pour le facteur sigma double », c'est-à-dire qu'ils peuvent répondre à deux facteurs sigma différents, comme indiqué dans RegulonDB. Les plus courants sont les promoteurs qui peuvent répondre à la fois à sigma 70 et à sigma 38 (illustré sur la figure). Des études sur la dynamique de ces gènes ont montré que lorsque les cellules entrent en croissance stationnaire, elles sont presque aussi induites que les gènes qui ont une préférence pour σ38 seul. Ce niveau d'induction s'est avéré prévisible à partir de leur séquence promotrice. Un modèle de leur dynamique est présenté sur la figure. À l'avenir, ces promoteurs pourraient devenir des outils utiles dans les constructions génétiques synthétiques chez E. coli .