L'ARN messager de transfert (abrégé ARNtm , également connu sous le nom d'ARN 10Sa et par son nom génétique SsrA ) est une molécule d'ARN bactérienne avec des propriétés de type ARNt et ARN messager . L'ARNtm forme un complexe ribonucléoprotéique ( tmRNP ) avec la petite protéine B (SmpB), le facteur d'élongation Tu ( EF-Tu ) et la protéine ribosomique S1. En trans -traduction, l'ARNtm et ses protéines associées se lient aux ribosomes bactériens qui se sont arrêtés au milieu de la biosynthèse des protéines , par exemple lorsqu'ils atteignent la fin d'un ARN messager qui a perdu son codon stop. L'ARNtm est remarquablement polyvalent : il recycle le ribosome arrêté, ajoute une étiquette induisant la protéolyse au polypeptide inachevé et facilite la dégradation de l' ARN messager aberrant . Dans la majorité des bactéries, ces fonctions sont assurées par des ARNtm monobloc standard. Chez d’autres espèces bactériennes, un gène ssrA permuté produit un ARNtm en deux parties dans lequel deux chaînes d’ARN distinctes sont jointes par appariement de bases.
Découverte et premiers travaux
L'ARNtm a été désigné pour la première fois ARN 10Sa en 1979, après qu'une fraction électrophorétique mixte « 10S » d' ARN d'Escherichia coli ait été résolue en ARNtm et en ARN RNase P de taille similaire (10Sb). La présence de pseudouridine dans l'ARN 10S mixte a suggéré que l'ARNtm possède des bases modifiées trouvées également dans l'ARNt . La similitude de l'extrémité 3' de l'ARNtm avec la tige-boucle T de l'ARNt a été reconnue pour la première fois lors du séquençage de ssrA de Mycobacterium tuberculosis . Une comparaison de séquence ultérieure a révélé le domaine de type ARNt complet (TLD) formé par les extrémités 5' et 3' de l'ARNtm, y compris la tige acceptrice avec des éléments comme ceux de l'ARNt d'alanine qui favorisent son aminoacylation par l'alanine-ARNt ligase . Il a également révélé des différences avec l'ARNt : le bras anticodon est manquant dans l'ARNtm et la région du bras D est une boucle sans paires de bases.
Structure
Structure secondaire des ARNm monoblocs standards

La structure secondaire complète de l'ARNtm d'E. coli a été élucidée par analyse comparative de séquence et sondage structurel . Les paires de bases Watson-Crick et GU ont été identifiées en comparant les séquences d'ARNtm bactériennes à l'aide de méthodes de calcul automatisées en combinaison avec des procédures d'alignement manuel . La figure ci-jointe montre le modèle d'appariement de bases de cet ARNtm prototypique, qui est organisé en 12 hélices supportées phylogénétiquement (également appelées appariements P1 à P12), certaines divisées en segments hélicoïdaux.
Une caractéristique importante de chaque ARNtm est le domaine de type ARNt conservé (TLD), composé des hélices 1, 12 et 2a (analogues de la tige acceptrice, de la tige T et de la tige variable de l'ARNt, respectivement), et contenant les extrémités 5' monophosphate et 3' CCA alanylatable. La région de type ARNm (MLR) est dans l'ARNtm standard une grande boucle contenant des pseudo-nœuds et une séquence codante (CDS) pour le peptide marqueur , marqué par le codon de reprise et le codon d'arrêt . Le peptide marqueur codé (ANDENYALAA dans E. coli ) varie selon les bactéries, peut-être en fonction de l'ensemble des protéases et des adaptateurs disponibles.
Les ARNtm contiennent généralement quatre pseudo-nœuds , un (pk1) en amont du CDS du peptide marqueur, et les trois autres pseudo-nœuds (pk2 à pk4) en aval du CDS. Les régions de pseudo-nœuds, bien que généralement conservées, sont plastiques au cours de l'évolution. Par exemple, dans les ARNtm (en une seule pièce) des cyanobactéries , pk4 est remplacé par deux pseudo-nœuds plus petits disposés en tandem. Cela suggère que le repliement de l'ARNtm en dehors du TLD peut être important, mais la région des pseudo-nœuds manque de résidus conservés et les pseudo-nœuds sont parmi les premières structures à être perdues lorsque les séquences ssrA divergent dans les lignées de plastides et d'endosymbiotes. L'appariement des bases dans la région des trois pseudo-nœuds de l'ARNtm d'E. coli est perturbé pendant la trans-traduction.
ARNtm en deux parties
La permutation circulaire de ssrA a été rapportée dans trois lignées principales : i) toutes les alphaprotéobactéries et les mitochondries primitives des protistes jacobides, ii) deux groupes disjoints de cyanobactéries ( Gloeobacter et un clade contenant Prochlorococcus et de nombreux Synechococcus ), et iii) certains membres des bêtaprotéobactéries ( Cupriavidus et certaines Rhodocyclales). Tous produisent la même forme globale en deux parties (pièces acceptrices et codantes), équivalente à la forme standard coupée en aval du cadre de lecture. Aucun ne conserve plus de deux pseudonœuds par rapport aux quatre (ou plus) de l'ARNtm standard.
Les alphaprotéobactéries ont deux séquences de signature : le remplacement de la séquence T-loop typique TΨCRANY par GGCRGUA et la séquence AACAGAA dans la grande boucle du pseudo-nœud 3'-terminal. Dans les mitochondries, le MLR a été perdu et une re-permutation remarquable de ssrA mitochondriale donne un petit produit d'un seul tenant dans Jakoba libera .
Les cyanobactéries constituent le cas le plus plausible d’évolution d’un gène permuté à partir d’un gène standard, en raison des similitudes de séquence remarquables entre les deux types de gènes tels qu’ils apparaissent dans différentes souches de Synechococcus .
Traitement de l'ARNtm
La plupart des ARNtm sont transcrits sous forme de précurseurs plus gros qui sont traités de la même manière que les ARNt . Le clivage à l'extrémité 5' se fait par la ribonucléase P. Plusieurs exonucléases peuvent participer au traitement de l'extrémité 3' des ARNtm, bien que la RNase T et la RNase PH soient les plus efficaces. Selon l'espèce bactérienne, le 3'-CCA est soit codé, soit ajouté par la nucléotidyltransférase de l'ARNt .
Un traitement similaire sur les sites internes du précurseur permuté de l'ARNtm explique sa division physique en deux parties. Les ARNtm en deux parties ont deux extrémités supplémentaires dont le traitement doit être pris en compte. Pour les alphaprotéobactéries, une extrémité 5' est le site de départ non traité de la transcription. L'extrémité 3' éloignée peut dans certains cas être le résultat d'une terminaison indépendante du rho.
Structures tridimensionnelles
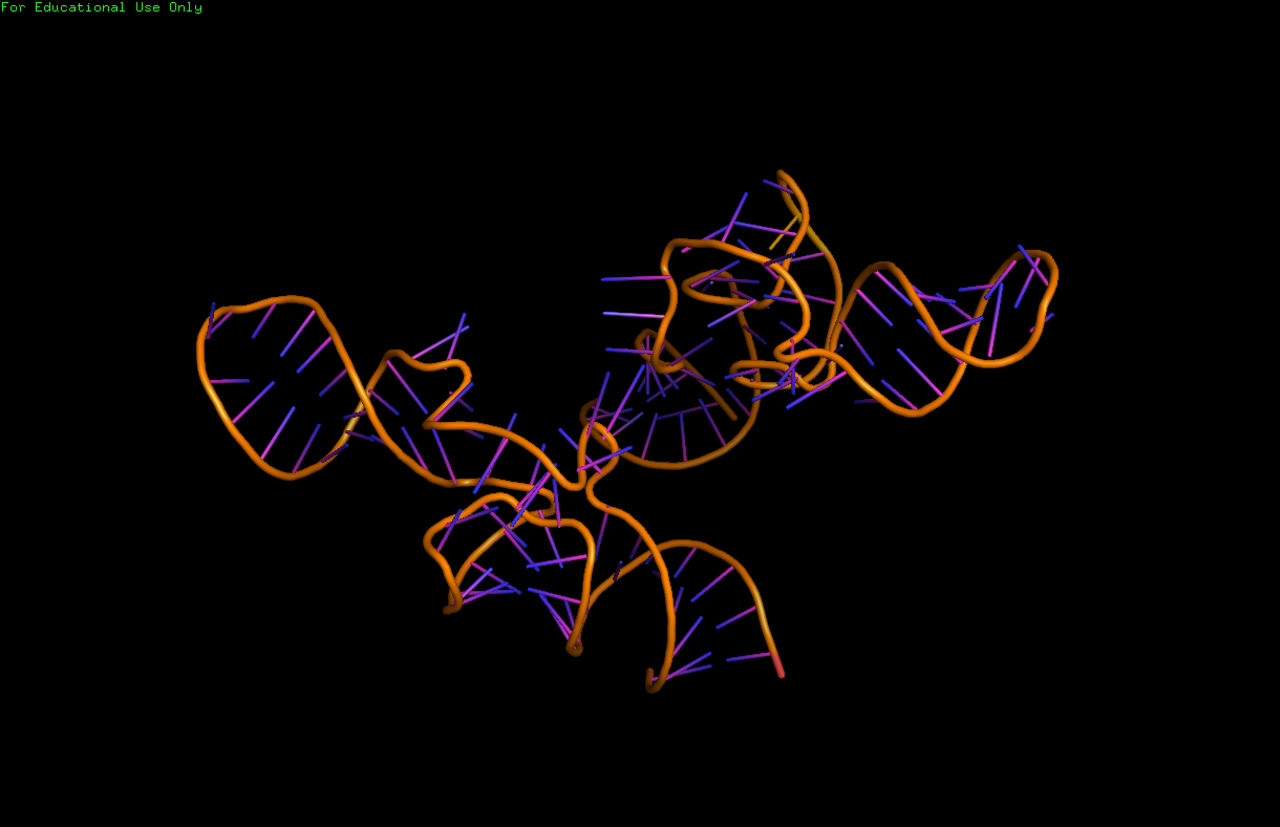
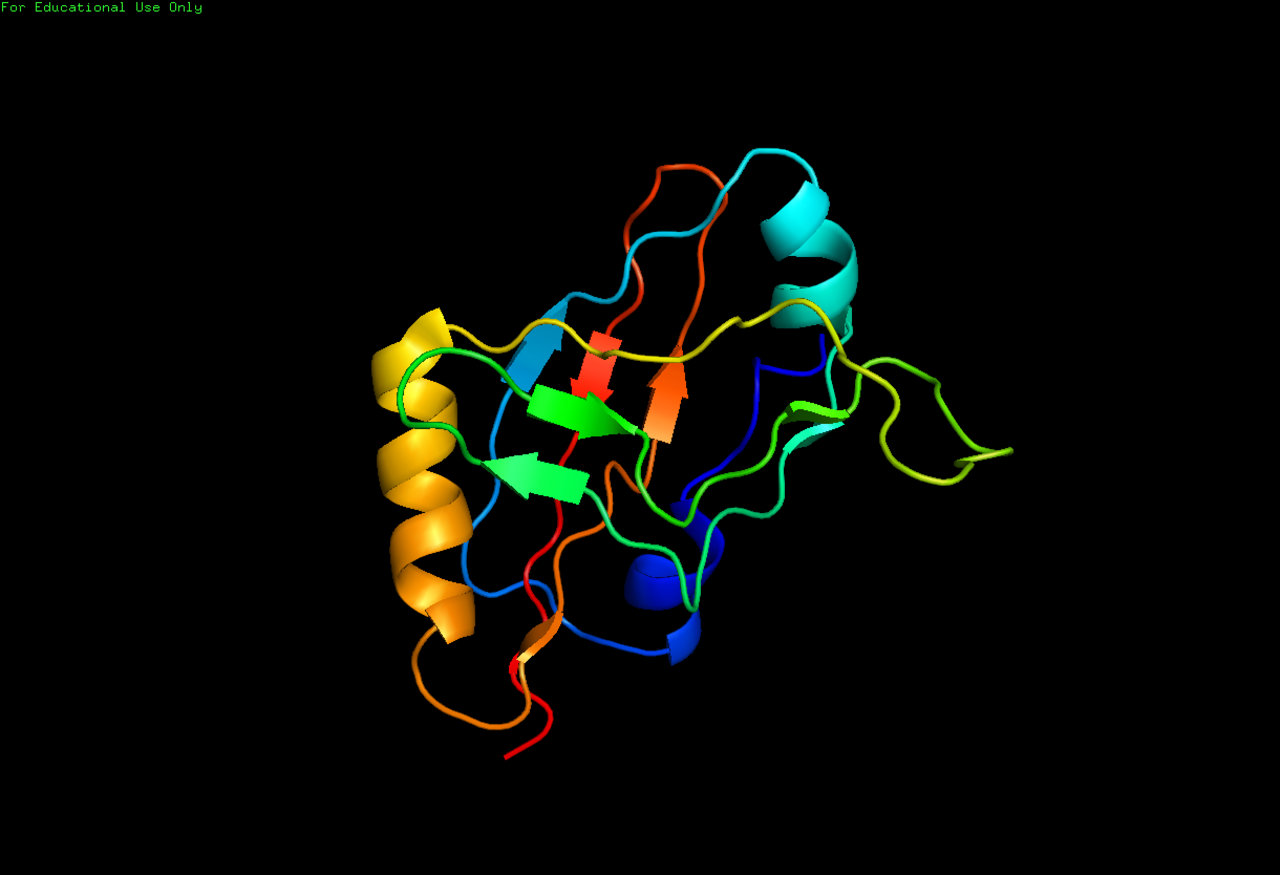
Les structures à haute résolution des molécules complètes d'ARNtm ne sont actuellement pas disponibles et peuvent être difficiles à obtenir en raison de la flexibilité inhérente de la MLR. En 2007, la structure cristalline du TLD de Thermus thermophilus lié à la protéine SmpB a été obtenue à une résolution de 3 Å. Cette structure montre que SmpB imite la tige D et l'anticodon d'un ARNt canonique tandis que la section hélicoïdale 2a de l'ARNtm correspond au bras variable de l'ARNt. Une étude par cryomicroscopie électronique de l'ARNtm à un stade précoce de la trans-traduction montre la relation spatiale entre le ribosome et le tmRNP (ARNtm lié à la protéine EF-Tu ). Le TLD est situé près du centre associé à la GTPase dans la sous-unité ribosomique 50S ; l'hélice 5 et les pseudo-nœuds pk2 à pk4 forment un arc autour du bec de la sous-unité ribosomique 30S.
Trans-traduction
Le codage par ARNtm a été découvert en 1995 lorsque Simpson et ses collègues ont surexprimé la cytokine de souris IL-6 dans E. coli et ont trouvé plusieurs peptides dérivés de cytokines tronqués , chacun marqué aux extrémités carboxyles avec la même extension de résidu de 11 acides aminés (A)ANDENYALAA. À l'exception de l' alanine N-terminale , qui provient de l'extrémité 3' de l'ARNtm lui-même, cette séquence de marquage a été tracée jusqu'à un court cadre de lecture ouvert dans l'ARNtm d'E. coli . Keiler et al. ont reconnu que le peptide marqueur confère la protéolyse et ont proposé le modèle de traduction trans pour l'action de l'ARNtm.
Bien que les détails du mécanisme de trans -traduction soient à l'étude, il est généralement admis que l'ARNtm occupe d'abord le site A vide du ribosome bloqué . Par la suite, le ribosome se déplace de l'extrémité 3' de l' ARN messager tronqué vers le codon de reprise du MLR, suivi d'une étape sujette au glissement à partir de laquelle la traduction continue normalement jusqu'à ce que le codon d'arrêt de l'ARNtm dans le cadre soit rencontré. La trans-traduction est essentielle chez certaines espèces bactériennes, tandis que d'autres bactéries ont besoin de l'ARNtm pour survivre lorsqu'elles sont soumises à des conditions de croissance stressantes. On pense que l'ARNtm peut aider la cellule à résister aux antibiotiques en sauvant les ribosomes bloqués par les antibiotiques. Selon l'organisme, le peptide marqueur peut être reconnu par une variété de protéases ou d'adaptateurs de protéase.
Éléments génétiques mobiles et gène tmRNA

ssrA est à la fois une cible pour certains ADN mobiles et un passager pour d'autres. Il a été découvert qu'il est interrompu par trois types d'éléments mobiles. Par différentes stratégies, aucun d'entre eux ne perturbe la fonction des gènes : les introns du groupe I s'éliminent par auto-épissage, les éléments palindromiques rickettsiaux (RPE) s'insèrent dans des sites inoffensifs et les îlots génomiques codant pour l'intégrase divisent leur cible ssrA mais restaurent la partie séparée.
Le gène ssrA non chromosomique a été détecté pour la première fois dans une étude génomique des mycobactériophages (dans 10 % des phages). D'autres éléments mobiles, notamment des plasmides et des îlots génomiques, ont été trouvés porteurs du gène ssrA . Un cas intéressant est celui de Rhodobacter sphaeroides ATCC 17025, dont le gène natif d'ARNtm est perturbé par un îlot génomique ; contrairement à tous les autres îlots génomiques dans les gènes d'ARNtm (ou d'ARNt), cet îlot a inactivé le gène cible natif sans restauration, mais compense en portant son propre gène d'ARNtm. Un parent très inhabituel du gène ssrA se trouve dans le mycobactériophage lytique DS6A, qui code un peu plus que le TLD.
ARNtm mitochondriaux (ssrAgène)
Une forme mitochondriale codée et structurellement réduite de tmRNA (mt-tmRNA) a été postulée pour la première fois pour le flagellé jacobide Reclinomonas americana . Par la suite, la présence d'un gène mitochondrial ( ssrA ) codant pour tmRNA, ainsi que des sites de transcription et de traitement de l'ARN ont été confirmés pour tous les membres des jacobides sauf un . Des preuves fonctionnelles, c'est-à-dire l'aminoacylation mt-tmRNA avec de l'alanine , sont disponibles pour Jakoba libera . Plus récemment, ssrA a également été identifié dans les génomes mitochondriaux des oomycètes . Comme dans les α-protéobactéries (les ancêtres des mitochondries ), les mt-tmRNA sont des molécules d'ARN en deux parties permutées de manière circulaire, sauf chez Jakoba libera où le gène est revenu au codage d'une conformation d'ARNt en une seule partie.
Identification dessrAdans les génomes mitochondriaux

Les gènes d'ARNtm mitochondriaux ont été initialement reconnus comme de courtes séquences conservées chez les jacobides et qui ont le potentiel de se replier dans une structure secondaire distincte de type ARNt. Avec la disponibilité de neuf séquences complètes d'ADNmt des jacobides , et un outil de recherche de covariance considérablement amélioré (Infernal ; ), un modèle de covariance a été développé sur la base des ARNtm mitochondriaux des jacobides , qui a également identifié les gènes mitochondriaux ssrA chez les oomycètes . À l'heure actuelle, un total de 34 ARNtm mt d'oomycètes ont été détectés dans six genres : Albugo , Bremia , Phytophthora , Pseudoperonospora , Pythium et Saprolegnia . Un modèle de covariance construit avec des séquences de jakobid et d'oomycètes est désormais disponible sur Rfam sous le nom de « mt-tmRNA ».
Structure de l'ARNmt-tm
L'ARNtm bactérien standard se compose d'un domaine de type ARNt(Ala) (permettant l'ajout d'une alanine non codée aux ARNm qui manquent de code d'arrêt) et d'un domaine de type ARNm codant pour une étiquette protéique qui destine le polypeptide à la protéolyse. Le domaine de type ARNm a été perdu dans les ARNtmmt. L'analyse comparative des séquences indique des caractéristiques typiques des ARNtmmt. La séquence primaire de la tige acceptrice aminoacyle est la plus conservée. Cette partie de la molécule a un résidu A invariable en position discriminatrice et une paire GU en position 3 (sauf dans S eculamonas ecuadoriensis , qui a une paire GC) ; cette position est le site de reconnaissance de l'ARNt synthase alanyl. P2 est une hélice de longueur variable (3 à 10 paires de bases) et correspond à la tige anticodon des ARNt, mais sans boucle anticodon (car elle n'est pas nécessaire à la fonction de l'ARNtm). P2 stabilise la structure de type ARNt, mais quatre nucléotides invariants chez les oomycètes et les jakobidés suggèrent une fonction supplémentaire, actuellement non identifiée. P3 a cinq paires de bases et correspond au bras T des ARNt, mais avec des nucléotides consensus différents à la fois dans la région appariée et dans la boucle. La séquence de la boucle T est conservée chez les oomycètes et les jakobidés , avec seulement quelques déviations (par exemple, Saprolegnia ferax ). Enfin, au lieu de la tige D de type ARNt avec une boucle D raccourcie de trois nucléotides caractéristique des ARNtm bactériens, les homologues mitochondriaux ont une boucle très variable de 5 à 14 nt de long. La séquence intermédiaire (Int.) des ARNtm-mt en deux parties est riche en A+U et de longueur irrégulière (4-34 nt). ). Pour les modèles de structure secondaire des ARNtm-mt en une et deux parties, voir la figure 1.
Traitement et expression de l'ARNmt-tm
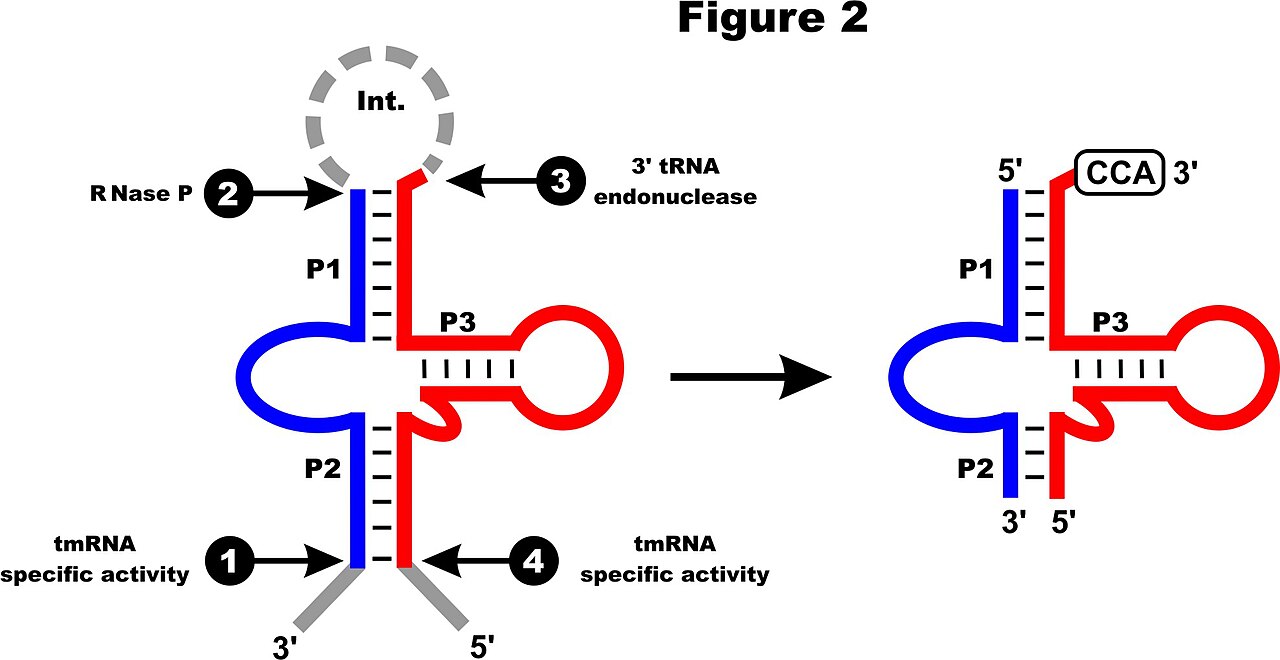
Les données RNA-Seq de Phytophthora sojae montrent un niveau d'expression similaire à celui des ARNt mitochondriaux voisins , et quatre sites de traitement majeurs confirment les extrémités prédites de l'ARNtm-mt mature. La molécule précurseur de l'ARNtm est probablement traitée par la RNase P et une endonucléase de traitement de l'ARNt 3' (voir la figure 2) ; cette dernière activité est supposée conduire à la suppression de la séquence intermédiaire. Après l'ajout de CCA au nucléotide discriminateur 3', l'ARNtm peut être chargé par l'alanyl-ARNt synthétase avec de l'alanine.