La sélection équilibrée fait référence à un certain nombre de processus sélectifs par lesquels plusieurs allèles (différentes versions d'un gène ) sont activement maintenus dans le pool génétique d'une population à des fréquences plus élevées que celles attendues par la seule dérive génétique . La sélection équilibrée est rare par rapport à la sélection purificatrice. Elle peut se produire par divers mécanismes, en particulier lorsque les hétérozygotes pour les allèles considérés ont une meilleure aptitude que l' homozygote . De cette façon, le polymorphisme génétique est conservé.
La preuve de l'existence d'une sélection équilibrée peut être trouvée dans le nombre d'allèles d'une population qui sont maintenus au-dessus des fréquences de taux de mutation. Toutes les recherches modernes ont montré que cette variation génétique significative est omniprésente dans les populations panmictiques .
Il existe plusieurs mécanismes (qui ne sont pas exclusifs à une population donnée) par lesquels la sélection équilibrée agit pour maintenir le polymorphisme. Les deux principaux mécanismes, les plus étudiés, sont l'avantage des hétérozygotes et la sélection dépendante de la fréquence.
Mécanismes
Avantage hétérozygote

Dans le cas de l'avantage hétérozygote , ou de la sélection hétérotique équilibrée , un individu hétérozygote à un locus génétique particulier a une meilleure condition physique qu'un individu homozygote. Les polymorphismes maintenus par ce mécanisme sont des polymorphismes équilibrés . En raison de fréquences élevées inattendues d'hétérozygotes et d'un niveau élevé de condition physique des hétérozygotes, l'avantage hétérozygote peut également être appelé « surdominance » dans certaines publications.
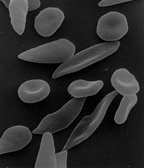
Un cas bien étudié est celui de l'anémie falciforme chez l'homme, une maladie héréditaire qui endommage les globules rouges . L'anémie falciforme est causée par l'hérédité d'un allèle (HgbS) du gène de l'hémoglobine des deux parents. Chez ces individus, l'hémoglobine des globules rouges est extrêmement sensible au manque d'oxygène, ce qui entraîne une espérance de vie plus courte. Une personne qui hérite du gène de la drépanocytose d'un parent et d'un allèle d'hémoglobine normal (HgbA) de l'autre, a une espérance de vie normale. Cependant, ces individus hétérozygotes, connus comme porteurs du trait drépanocytaire , peuvent souffrir de problèmes de temps à autre.
L'hétérozygote est résistant au parasite du paludisme qui tue un grand nombre de personnes chaque année. Il s'agit d'un exemple de sélection équilibrée entre la sélection féroce contre les drépanocytaires homozygotes et la sélection contre les homozygotes HgbA standard du paludisme. L'hétérozygote a un avantage permanent (une meilleure aptitude) partout où le paludisme existe. Le maintien de l'allèle HgbS par sélection positive est soutenu par des preuves significatives selon lesquelles les hétérozygotes ont une aptitude réduite dans les régions où le paludisme n'est pas répandu. Au Suriname, par exemple, l'allèle est maintenu dans les pools génétiques des descendants d'esclaves africains, car le Suriname souffre d'épidémies de paludisme pérennes. Curaçao, cependant, qui compte également une population importante d'individus descendants d'esclaves africains, n'est pas touché par le paludisme généralisé et n'a donc pas non plus la pression sélective nécessaire pour maintenir l'allèle HgbS. À Curaçao, l’allèle HgbS a diminué en fréquence au cours des 300 dernières années et finira par disparaître du pool génétique en raison du désavantage des hétérozygotes .
Sélection dépendante de la fréquence
Français La sélection dépendante de la fréquence se produit lorsque la fitness d'un phénotype dépend de sa fréquence par rapport aux autres phénotypes d'une population donnée. Dans la sélection dépendante de la fréquence positive, la fitness d'un phénotype augmente à mesure qu'il devient plus courant. Dans la sélection dépendante de la fréquence négative , la fitness d'un phénotype diminue à mesure qu'il devient plus courant. Par exemple, dans le changement de proie , les formes rares de proies sont en fait plus adaptées en raison des prédateurs qui se concentrent sur les formes les plus fréquentes. Comme la prédation fait baisser les fréquences démographiques de la forme commune de proie, la forme autrefois rare de proie devient la forme la plus courante. Ainsi, la forme avantageuse est désormais la forme désavantageuse. Cela peut conduire à des cycles d'expansion et de récession des formes de proies. Les interactions hôte-parasite peuvent également conduire à une sélection dépendante de la fréquence négative, conformément à l'hypothèse de la Reine rouge. Par exemple, le parasitisme de l'escargot d'eau douce de Nouvelle-Zélande ( Potamopyrgus antipodarum ) par le trématode Microphallus sp. Les résultats de cette étude sont une diminution des fréquences des génotypes les plus fréquemment hébergés sur plusieurs générations. Plus un génotype est commun au cours d'une génération, plus il est vulnérable au parasitisme par Microphallus sp. [ Il convient de noter que dans ces exemples, aucun morphe phénotypique, ni aucun génotype ne sont entièrement éteints d'une population, et aucun morphe phénotypique ni aucun génotype ne sont sélectionnés pour la fixation. Ainsi, le polymorphisme est maintenu par une sélection dépendante de la fréquence négative.
La condition physique varie dans le temps et dans l'espace
La fitness d'un génotype peut varier considérablement entre les stades larvaire et adulte, ou entre les différentes parties d'un habitat. La variation dans le temps, contrairement à la variation dans l'espace, n'est pas suffisante en soi pour maintenir plusieurs types, car en général, le type avec la fitness moyenne géométrique la plus élevée prendra le dessus, mais il existe un certain nombre de mécanismes qui rendent possible une coexistence stable.
Exemples plus complexes
Les espèces dans leur habitat naturel sont souvent beaucoup plus complexes que les exemples typiques des manuels scolaires.
Escargot des bois
L'escargot de forêt, Cepaea nemoralis , est célèbre pour le riche polymorphisme de sa coquille. Le système est contrôlé par une série d' allèles multiples . L'absence de bandes est le trait dominant le plus élevé, et les formes de bandes sont contrôlées par des gènes modificateurs (voir épistasie ).

En Angleterre, l'escargot est régulièrement la proie de la grive musicienne Turdus philomelos , qui l'ouvre sur des enclumes (grosses pierres). Les fragments s'accumulent alors, ce qui permet aux chercheurs d'analyser les escargots capturés. Les grives chassent à vue et capturent sélectivement les formes qui correspondent le moins bien à l'habitat . Les colonies d'escargots se trouvent dans les bois, les haies et les prairies, et la prédation détermine la proportion de phénotypes (morphes) trouvés dans chaque colonie.

Un deuxième type de sélection s'opère également chez l'escargot, certains hétérozygotes ayant un avantage physiologique sur les homozygotes. Troisièmement, une sélection apostatique est probable, les oiseaux choisissant préférentiellement la forme la plus commune. Il s'agit de l'effet de « modèle de recherche », où un prédateur à prédominance visuelle persiste à cibler la forme qui a donné un bon résultat, même si d'autres formes sont disponibles.
Le polymorphisme survit dans presque tous les habitats, bien que les proportions de morphes varient considérablement. Les allèles contrôlant le polymorphisme forment un supergène dont la liaison est si étroite qu'elle est presque absolue. Ce contrôle préserve la population d'une forte proportion de recombinants indésirables.
Chez cette espèce, la prédation par les oiseaux semble être la principale (mais pas la seule) force sélective à l'origine du polymorphisme. Les escargots vivent sur des milieux hétérogènes et les grives sont habiles à détecter les correspondances médiocres. L'hérédité de la diversité physiologique et cryptique est également préservée par l'avantage hétérozygote du supergène. Des travaux récents ont inclus l'effet de la couleur de la coquille sur la thermorégulation, et une sélection plus large d'influences génétiques possibles est également envisagée.
Polymorphisme chromosomique chezDrosophile
Dans les années 1930, Theodosius Dobzhansky et ses collègues ont collecté Drosophila pseudoobscura et D. persimilis dans des populations sauvages de Californie et des États voisins. En utilisant la technique de Painter , ils ont étudié les chromosomes polytènes et ont découvert que toutes les populations sauvages étaient polymorphes pour les inversions chromosomiques . Toutes les mouches se ressemblent, quelles que soient les inversions qu'elles portent, ce qui constitue un exemple de polymorphisme cryptique. Les preuves se sont accumulées pour montrer que la sélection naturelle en était responsable :
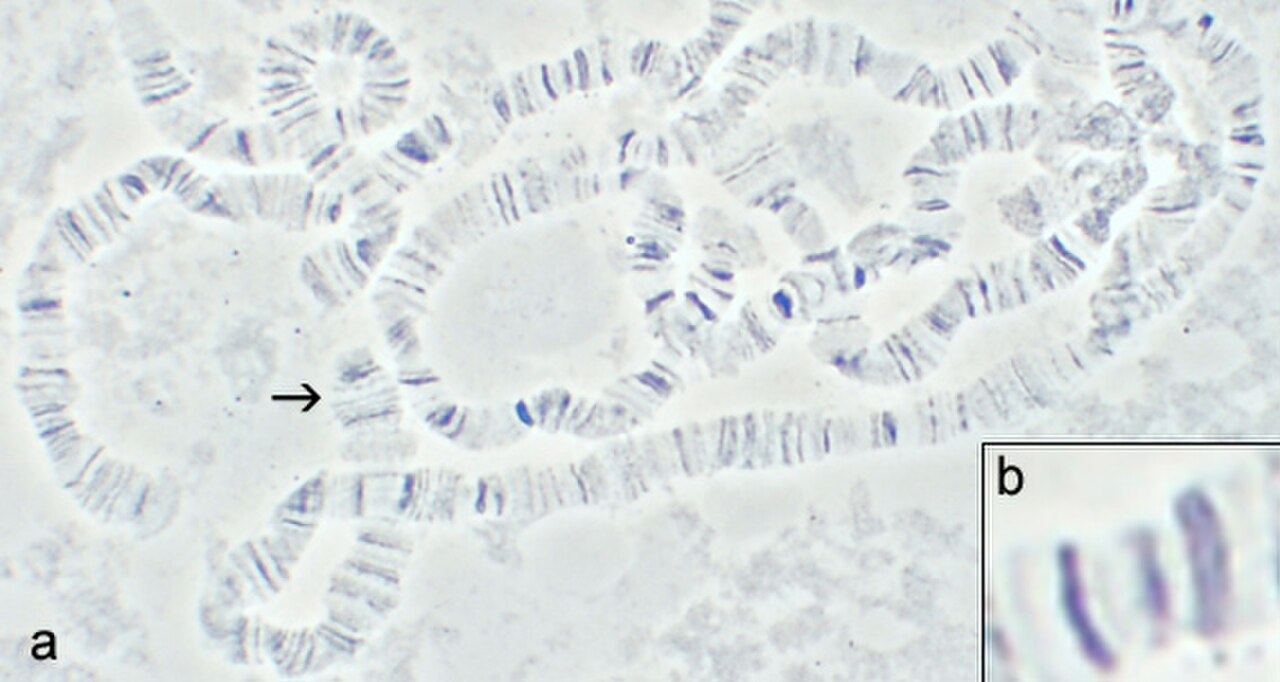
- Les valeurs des inversions hétérozygotes du troisième chromosome étaient souvent bien plus élevées qu'elles n'auraient dû l'être sous l'hypothèse nulle : s'il n'y a aucun avantage pour aucune forme, le nombre d'hétérozygotes devrait être conforme à N s (nombre dans l'échantillon) = p 2 + 2pq + q 2 où 2pq est le nombre d'hétérozygotes (voir équilibre de Hardy–Weinberg ).
- En utilisant une méthode inventée par L'Heretier et Teissier, Dobzhansky a élevé des populations dans des cages de population , ce qui permettait de nourrir, de reproduire et d'échantillonner tout en empêchant les évasions. Cette méthode avait l'avantage d'éliminer la migration comme explication possible des résultats. Les stocks contenant des inversions à une fréquence initiale connue peuvent être maintenus dans des conditions contrôlées. Il a été constaté que les différents types de chromosomes ne fluctuent pas au hasard, comme ils le feraient s'ils étaient sélectivement neutres, mais s'ajustent à certaines fréquences auxquelles ils se stabilisent.
- Des proportions différentes de morphes chromosomiques ont été trouvées dans différentes zones. Il existe, par exemple, un cline polymorph-ratio chez D. robusta le long d'un transect de 29 km (18 miles) près de Gatlinburg , TN, passant de 300 m (1 000 pieds) à 1 200 m (4 000 pieds). De plus, les mêmes zones échantillonnées à différentes périodes de l'année ont donné lieu à des différences significatives dans les proportions des formes. Cela indique un cycle régulier de changements qui ajustent la population aux conditions saisonnières. Pour ces résultats, la sélection est de loin l'explication la plus probable.
- Enfin, les morphes ne peuvent pas être maintenus aux niveaux élevés trouvés simplement par mutation, et la dérive n'est pas une explication possible lorsque les effectifs de population sont élevés.
En 1951, Dobzhansky était persuadé que les morphes chromosomiques étaient maintenus dans la population grâce à l'avantage sélectif des hétérozygotes, comme c'est le cas pour la plupart des polymorphismes.