La classification de Baltimore est un système utilisé pour classer les virus en fonction de leur mode de synthèse de l'ARN messager (ARNm). En organisant les virus en fonction de leur mode de production d'ARNm, il est possible d'étudier les virus qui se comportent de manière similaire en tant que groupe distinct. Sept groupes de Baltimore sont décrits qui prennent en compte si le génome viral est constitué d' acide désoxyribonucléique (ADN) ou d'acide ribonucléique (ARN), si le génome est monocaténaire ou bicaténaire et si le sens d'un génome d'ARN monocaténaire est positif ou négatif.
La classification de Baltimore correspond également étroitement à la manière dont le génome est répliqué , de sorte que la classification de Baltimore est utile pour regrouper les virus pour la transcription et la réplication. Certains sujets relatifs aux virus sont associés à plusieurs groupes de Baltimore spécifiques, tels que des formes spécifiques de traduction de l'ARNm et la gamme d'hôtes de différents types de virus. Les caractéristiques structurelles telles que la forme de la capside virale , qui stocke le génome viral, et l'histoire évolutive des virus ne sont pas nécessairement liées aux groupes de Baltimore.
La classification de Baltimore a été créée en 1971 par le virologue David Baltimore . Depuis lors, il est devenu courant parmi les virologues d'utiliser la classification de Baltimore parallèlement à la taxonomie standard des virus, qui est basée sur l'histoire évolutive. En 2018 et 2019, la classification de Baltimore a été partiellement intégrée à la taxonomie des virus sur la base de preuves que certains groupes descendaient d'ancêtres communs. Divers domaines, règnes et phylums correspondent désormais à des groupes de Baltimore spécifiques.
Aperçu
La classification de Baltimore regroupe les virus en fonction de leur mode de synthèse de l'ARNm. Les caractéristiques directement liées à cela incluent le fait que le génome soit constitué d' acide désoxyribonucléique (ADN) ou d'acide ribonucléique (ARN), le brin du génome, qui peut être simple ou double brin, et le sens d'un génome simple brin, qui est soit positif soit négatif. Le principal avantage de la classification de Baltimore est qu'en classant les virus selon les caractéristiques susmentionnées, les virus qui se comportent de la même manière peuvent être étudiés en tant que groupes distincts. Il existe sept groupes de Baltimore numérotés avec des chiffres romains, énumérés ci-après.
- Groupe I : virus à ADN double brin
- Groupe II : virus à ADN simple brin
- Groupe III : virus à ARN double brin
- Groupe IV : virus à ARN simple brin à polarité positive
- Groupe V : virus à ARN simple brin à polarité négative
- Groupe VI : virus à ARN simple brin avec un ADN intermédiaire dans leur cycle de vie
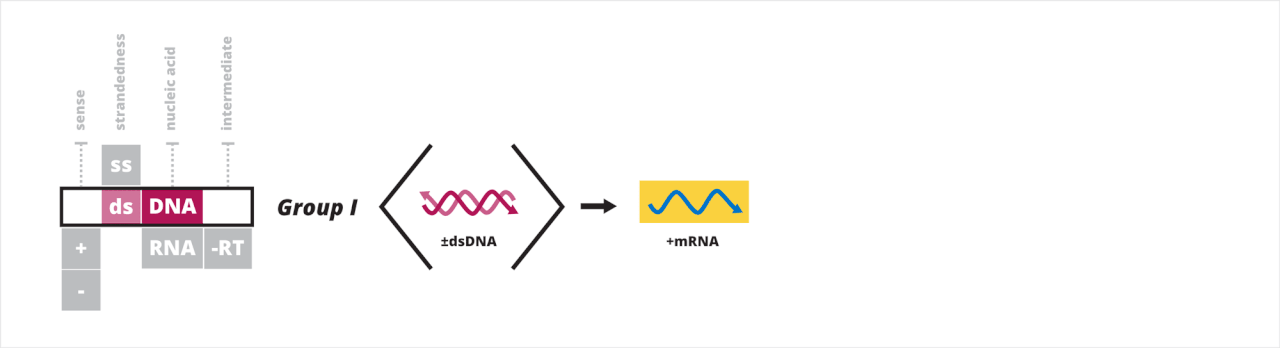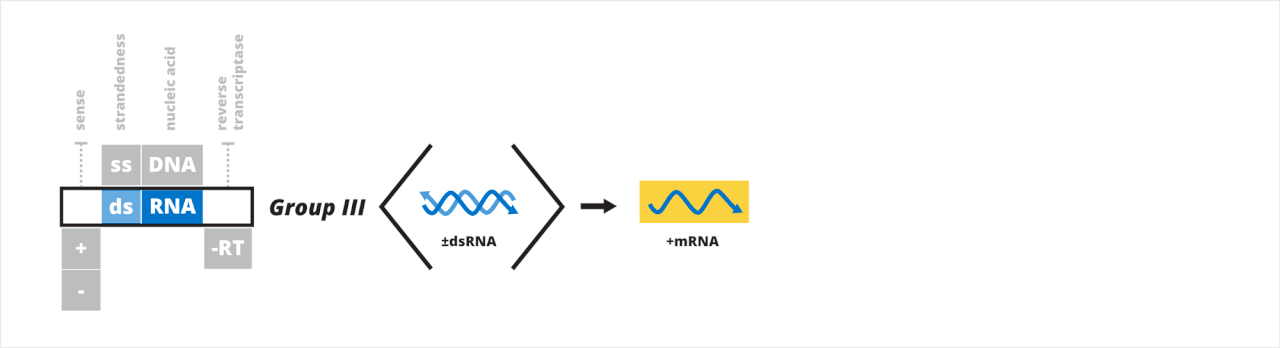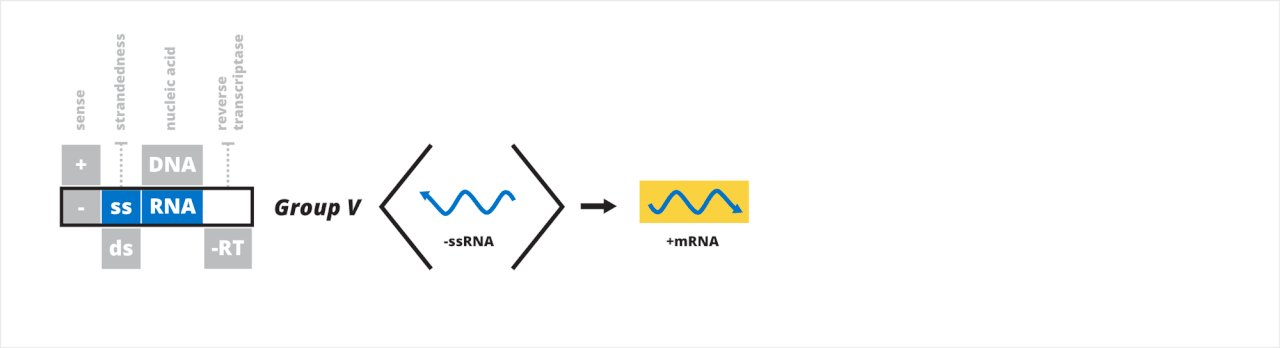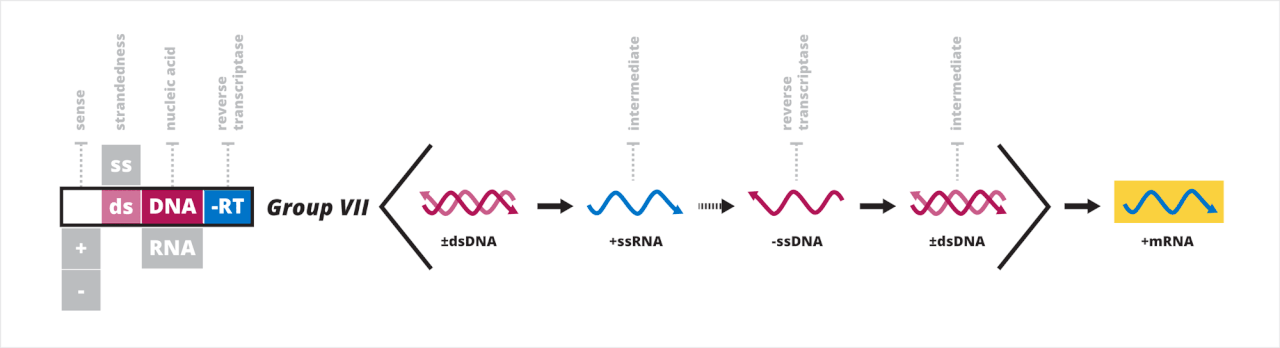
- Groupe VII : virus à ADN double brin avec un intermédiaire ARN dans leur cycle biologique
La classification de Baltimore est principalement basée sur la transcription du génome viral, et les virus de chaque groupe partagent généralement les mêmes modes de synthèse de l'ARNm. Bien que ce ne soit pas l'objectif direct de la classification de Baltimore, les groupes sont organisés de telle manière que les virus de chaque groupe ont également généralement les mêmes mécanismes de réplication du génome viral. Pour cette raison, la classification de Baltimore fournit des informations sur les parties transcription et réplication du cycle de vie viral . Les caractéristiques structurelles d'une particule virale, appelée virion, telles que la forme de la capside virale et la présence d'une enveloppe virale , une membrane lipidique qui entoure la capside, n'ont pas de relation directe avec les groupes de Baltimore, et les groupes ne présentent pas nécessairement de relation génétique basée sur l'histoire évolutive.
Classification
Virus à ADN
Les virus à ADN ont un génome composé d'acide désoxyribonucléique (ADN) et sont organisés en deux groupes : les virus à ADN double brin (ADNdb) et les virus à ADN simple brin (ADNsb). Ils sont classés en quatre domaines distincts : Adnaviria , Duplodnaviria , Monodnaviria et Varidnaviria . Beaucoup n'ont pas encore été classés dans un domaine.
Groupe I : virus à ADN double brin
Le premier groupe de Baltimore contient des virus qui ont un génome à ADN double brin (ADNdb). Tous les virus à ADNdb ont leur ARNm synthétisé selon un processus en trois étapes. Tout d'abord, un complexe de préinitiation de la transcription se lie à l'ADN en amont du site où la transcription commence, permettant le recrutement d'une ARN polymérase hôte . Ensuite, une fois l'ARN polymérase recrutée, elle utilise le brin négatif comme modèle pour synthétiser les brins d'ARNm. Troisièmement, l'ARN polymérase termine la transcription lorsqu'elle atteint un signal spécifique, tel qu'un site de polyadénylation .
Les virus à ADN double brin utilisent plusieurs mécanismes pour répliquer leur génome. La réplication bidirectionnelle, dans laquelle deux fourches de réplication sont établies sur un site d'origine de réplication et se déplacent dans des directions opposées, est largement utilisée. Un mécanisme de cercle roulant qui produit des brins linéaires tout en progressant dans une boucle autour du génome circulaire est également courant. Certains virus à ADN double brin utilisent une méthode de déplacement de brin par laquelle un brin est synthétisé à partir d'un brin modèle, et un brin complémentaire est ensuite synthétisé à partir du brin synthétisé précédemment, formant un génome à ADN double brin. Enfin, certains virus à ADN double brin sont répliqués dans le cadre d'un processus appelé transposition réplicative par lequel un génome viral dans l'ADN d'une cellule hôte est répliqué dans une autre partie d'un génome hôte.
Les virus à ADN double brin peuvent être subdivisés en virus qui se répliquent dans le noyau, et qui sont donc relativement dépendants de la machinerie de la cellule hôte pour la transcription et la réplication, et ceux qui se répliquent dans le cytoplasme, auquel cas ils ont évolué ou acquis leurs propres moyens d'exécuter la transcription et la réplication. Les virus à ADN double brin sont également communément divisés en virus à ADN double brin à queue, faisant référence aux membres du domaine Duplodnaviria , généralement les bactériophages à queue de l'ordre Caudovirales , et les virus à ADN double brin sans queue ou sans queue du domaine Varidnaviria .
Les virus à ADN double brin sont classés dans trois des quatre domaines et comprennent de nombreux taxons qui ne sont assignés à aucun domaine :
- Tous les virus d' Adnaviria sont des virus à ADN double brin. Les virus de ce domaine infectent les archées .
- Tous les virus de la famille Duplodnaviria sont des virus à ADN double brin. Les virus de ce genre appartiennent à deux groupes : les bactériophages à queue chez les Caudovirales et les herpèsvirus chez les Herpesvirales .
- Chez Monodnaviria , les membres de la classe Papovaviricetes sont des virus à ADN double brin. Les virus de la classe Papovaviricetes constituent deux groupes : les papillomavirus et les polyomavirus .
- Tous les virus de la famille Varidnaviria sont des virus à ADN double brin. Les virus de ce genre comprennent les adénovirus , les virus géants et les poxvirus .
- Les taxons suivants qui ne sont pas assignés à un domaine contiennent exclusivement des virus à ADN double brin :
- Classes : Naldaviricetes
- Familles : Ampullaviridae , Bicaudaviridae , Clavaviridae , Fuselloviridae , Globuloviridae , Guttaviridae , Halspiviridae , Ovaliviridae , Plasmaviridae , Polydnaviridae , Portogloboviridae , Thaspiviridae
- Genres : Dinodnavirus , Rhizidiovirus
Groupe II : virus à ADN simple brin
Le deuxième groupe de Baltimore contient des virus qui ont un génome à ADN simple brin (ADNss). Les virus à ADNss ont le même mode de transcription que les virus à ADNds. Cependant, comme le génome est simple brin, il est d'abord transformé en une forme double brin par une ADN polymérase lors de son entrée dans une cellule hôte. L'ARNm est ensuite synthétisé à partir de la forme double brin. La forme double brin des virus à ADNss peut être produite soit directement après l'entrée dans une cellule, soit à la suite de la réplication du génome viral. Les virus à ADNss eucaryotes sont répliqués dans le noyau.
La plupart des virus à ADN simple brin contiennent des génomes circulaires qui sont répliqués via la réplication en cercle roulant (RCR). La réplication en cercle roulant de l'ADN simple brin est initiée par une endonucléase qui se lie au brin positif et le clive, permettant à une ADN polymérase d'utiliser le brin négatif comme modèle pour la réplication. La réplication progresse dans une boucle autour du génome en étendant l'extrémité 3' du brin positif, en déplaçant le brin positif précédent, et l'endonucléase clive à nouveau le brin positif pour créer un génome autonome qui est ligaturé dans une boucle circulaire. Le nouvel ADN simple brin peut être conditionné dans des virions ou répliqué par une ADN polymérase pour former une forme double brin pour la transcription ou la poursuite du cycle de réplication.
Les parvovirus contiennent des génomes d'ADN simple brin linéaires qui sont répliqués via la réplication en épingle à cheveux roulante (RHR), qui est similaire à la RCR. Les génomes des parvovirus ont des boucles en épingle à cheveux à chaque extrémité du génome qui se déplient et se replient de manière répétée pendant la réplication pour changer la direction de la synthèse de l'ADN afin de se déplacer d'avant en arrière le long du génome, produisant de nombreuses copies du génome dans un processus continu. Les génomes individuels sont ensuite excisés de cette molécule par l'endonucléase virale. Pour les parvovirus, le brin sens positif ou négatif peut être emballé dans des capsides, variant d'un virus à l'autre.
Presque tous les virus à ADN simple brin ont un génome à sens positif, mais il existe quelques exceptions et particularités. La famille des Anelloviridae est la seule famille d'ADN simple brin dont les membres ont un génome à sens négatif, qui est circulaire. Les parvovirus, comme mentionné précédemment, peuvent conditionner le brin positif ou négatif dans les virions. Enfin, les bidnavirus conditionnent à la fois les brins linéaires positifs et négatifs. Dans tous les cas, le sens des virus à ADN simple brin, contrairement aux virus à ARN simple brin, n'est pas suffisant pour séparer les virus à ADN simple brin en deux groupes puisque tous les génomes viraux à ADN simple brin sont convertis en formes d'ADN double brin avant la transcription et la réplication.
Les virus à ADN simple brin sont classés dans l'un des quatre domaines et comprennent plusieurs familles qui ne sont assignées à aucun domaine :
- Chez Monodnaviria , tous les membres, à l'exception des virus de Papovaviricetes, sont des virus à ADN simple brin.
- Les familles non attribuées Anelloviridae et Spiraviridae sont des familles de virus à ADN simple brin.
- Les virus de la famille Finnlakeviridae contiennent des génomes à ADN simple brin. Finnlakeviridae n'est pas assigné à un domaine mais est un membre proposé de Varidnaviria .
Virus à ARN
Les virus à ARN ont un génome composé d'acide ribonucléique (ARN) et comprennent trois groupes : les virus à ARN double brin (ARNdb), les virus à ARN simple brin positif (+ARNss) et les virus à ARN simple brin négatif (-ARNss). La majorité des virus à ARN sont classés dans le règne des Orthornavirae, dans le domaine des riboviries . Les exceptions sont généralement les viroïdes et autres agents sous-viraux . Certains virus de cette dernière catégorie, comme le virus de l'hépatite D , sont classés dans le domaine des ribozyviries .
Groupe III : virus à ARN double brin
Le troisième groupe de Baltimore contient des virus qui ont un génome à ARN double brin (ARNdb). Après avoir pénétré dans une cellule hôte, le génome à ARNdb est transcrit en ARNm à partir du brin négatif par l'ARN polymérase virale dépendante de l'ARN (RdRp). L'ARNm peut être utilisé pour la traduction ou la réplication. L'ARNm simple brin est répliqué pour former le génome à ARNdb. L'extrémité 5' du génome peut être nue, coiffée ou liée de manière covalente à une protéine virale.
L'ARNdb n'est pas une molécule fabriquée par les cellules, c'est pourquoi la vie cellulaire a développé des systèmes antiviraux pour détecter et inactiver l'ARNdb viral. Pour contrer ce phénomène, de nombreux génomes d'ARNdb sont construits à l'intérieur de capsides, évitant ainsi d'être détectés à l'intérieur du cytoplasme de la cellule hôte. L'ARNm est expulsé de la capside afin d'être traduit ou d'être transloqué d'une capside mature à une capside progéniture. Alors que les virus à ARNdb ont généralement des capsides, les virus des familles Amalgaviridae et Endornaviridae n'ont pas été observés en train de former des virions et, par conséquent, ils semblent dépourvus de capsides. Les endornavirus sont également inhabituels dans la mesure où, contrairement aux autres virus à ARN, ils possèdent un seul et long cadre de lecture ouvert (ORF), ou partie traduisible, et une entaille spécifique au site dans la région 5' du brin positif.
Les virus à ARNdb sont classés en deux phylums au sein du royaume Orthornavirae du royaume Riboviria :
- Tous les virus de Duplornaviricota sont des virus à ARNdb.
- Chez Pisuviricota , tous les membres de la classe Duplopiviricetes sont des virus à ARNdb.
Groupe IV : virus à ARN simple brin à polarité positive

Le quatrième groupe de Baltimore contient des virus qui ont un génome à ARN simple brin positif (+ ARNss) de sens positif. Pour les virus à ARNss +, le génome fonctionne comme un ARNm, donc aucune transcription n'est nécessaire pour la traduction. Cependant, les virus à ARNss + produiront également des copies positives du génome à partir de brins négatifs d'un génome à ARNds intermédiaire. Cela agit à la fois comme un processus de transcription et de réplication puisque l'ARN répliqué est également un ARNm. L'extrémité 5' peut être nue, coiffée ou liée de manière covalente à une protéine virale, et l'extrémité 3' peut être nue ou polyadénylée.
De nombreux virus à ARN simple brin ne peuvent transcrire qu'une partie de leur génome. En général, les brins d'ARN sous-génomique (ARN simple brin) sont utilisés pour la traduction des protéines structurelles et de mouvement nécessaires aux stades intermédiaires et tardifs de l'infection. La transcription de l'ARN simple brin peut se produire en commençant la synthèse de l'ARN au sein du génome plutôt qu'à partir de l'extrémité 5', en arrêtant la synthèse de l'ARN au niveau de séquences spécifiques du génome ou, dans le cadre des deux méthodes précédentes, en synthétisant des séquences leaders à partir de l'ARN viral qui sont ensuite attachées aux brins d'ARN simple brin. Étant donné que la réplication est nécessaire à la synthèse de l'ARN simple brin, RdRp est toujours traduit en premier.
Étant donné que le processus de réplication du génome viral produit des molécules d'ARNdb intermédiaires, les virus à ARNss+ peuvent être ciblés par le système immunitaire de la cellule hôte. Pour éviter d'être détectés, les virus à ARNss+ se répliquent dans des vésicules associées à la membrane qui sont utilisées comme usines de réplication. À partir de là, seul l'ARNss+ viral, qui peut être de l'ARNm, pénètre dans la zone cytoplasmique principale de la cellule.
Les virus à ARN simple brin peuvent être subdivisés en ceux qui ont un ARNm polycistronique, qui code une polyprotéine qui est clivée pour former plusieurs protéines matures, et ceux qui produisent des ARNm sous-génomiques et subissent donc deux ou plusieurs cycles de traduction. Les virus à ARN simple brin sont inclus dans trois phylums du royaume des Orthornavirae dans le royaume des Riboviria :
- Tous les virus de Lenarviricota sont des virus à ARN simple brin.
- Tous les virus de Pisuviricota sont des virus à ARN simple brin, à l'exception de la classe Duplopiviricetes , dont les membres ont des génomes à ARN double brin.
- Tous les virus de Kitrinoviricota sont des virus à ARN simple brin.
Groupe V : virus à ARN simple brin à polarité négative
Le cinquième groupe de Baltimore contient des virus qui ont un génome à ARN simple brin (-ssRNA) de sens négatif. L'ARNm, qui est de sens positif, est transcrit directement à partir du génome de sens négatif. Le premier processus de transcription de l'-ssRNA implique la liaison de RdRp à une séquence leader à l'extrémité 3' du génome, la transcription d'un ARN leader 5' triphosphate qui est coiffé, puis l'arrêt et le redémarrage sur un signal de transcription qui est coiffé , continuant jusqu'à ce qu'un signal d'arrêt soit atteint. La deuxième manière est similaire mais au lieu de synthétiser une coiffe, RdRp peut utiliser le cap snatching , par lequel une courte séquence d'ARNm de la cellule hôte est prise et utilisée comme coiffe 5' de l'ARNm viral. L'-ssRNA génomique est répliqué à partir de l'antigénome de sens positif d'une manière similaire à la transcription, sauf en sens inverse en utilisant l'antigénome comme modèle pour le génome. RdRp se déplace de l'extrémité 3' à l'extrémité 5' de l'antigénome et ignore tous les signaux de transcription lors de la synthèse de l'ARNss génomique.
Différents virus à ARN simple brin utilisent des mécanismes spéciaux pour la transcription. La manière de produire la queue polyA peut être via le bégaiement de la polymérase , au cours duquel RdRp transcrit une adénine à partir de l'uracile , puis revient dans la séquence d'ARN avec l'ARNm pour le transcrire à nouveau, en continuant ce processus plusieurs fois jusqu'à ce que des centaines d'adénines aient été ajoutées à l'extrémité 3' de l'ARNm. De plus, certains virus à ARN simple brin sont ambisens, car les brins positifs et négatifs codent séparément les protéines virales, et ces virus produisent deux brins d'ARNm distincts : l'un directement à partir du génome et l'autre à partir d'un brin complémentaire.
Les virus à ARN simple brin peuvent être subdivisés de manière informelle entre ceux qui ont des génomes non segmentés et segmentés. Les virus à ARN simple brin non segmentés se répliquent dans le cytoplasme et les virus à ARN simple brin segmentés se répliquent dans le noyau. Pendant la transcription, le RdRp produit un brin d'ARNm monocistronique à partir de chaque segment du génome. Tous les virus à ARN simple brin sont classés dans le phylum Negarnaviricota dans le royaume Orthornavirae dans le royaume Riboviria . Negarnaviricota ne contient que des virus à ARN simple brin, donc « virus à ARN simple brin » est synonyme de Negarnaviricota . Negarnaviricota est divisé en deux sous-phylums : Haploviricotina , dont les membres synthétisent une structure de coiffe sur l'ARNm viral nécessaire à la synthèse des protéines, et Polyploviricotina , dont les membres obtiennent plutôt des coiffes sur l'ARNm via l'arrachage de coiffe.
Virus à transcription inverse
Les virus à transcription inverse (RT) ont des génomes composés d'ADN ou d'ARN et se répliquent par transcription inverse. Il existe deux groupes de virus à transcription inverse : les virus à ARN simple brin-RT (ssRNA-RT) et les virus à ADN double brin-RT (dsDNA-RT). Les virus à transcription inverse sont classés dans le règne des Pararnavirae dans le règne des Riboviria .
Groupe VI : virus à ARN simple brin avec un intermédiaire ADN
Le sixième groupe de Baltimore contient des virus qui ont un génome à ARN simple brin (sens positif) qui a un intermédiaire ADN ((+)ssRNA-RT) dans son cycle de réplication. Les virus à ARN simple brin-RT sont transcrits de la même manière que les virus à ADN, mais leurs génomes linéaires sont d'abord convertis en une forme dsADN par un processus appelé transcription inverse . L'enzyme virale transcriptase inverse synthétise un brin d'ADN à partir du brin dsARN, et le brin d'ARN est dégradé et remplacé par un brin d'ADN pour créer un génome dsADN. Le génome est ensuite intégré dans l'ADN de la cellule hôte, où il est maintenant appelé provirus . L' ARN polymérase II de la cellule hôte transcrit ensuite l'ARN dans le noyau à partir de l'ADN proviral. Une partie de cet ARN peut devenir de l'ARNm tandis que d'autres brins deviendront des copies du génome viral pour la réplication.
Les virus ssRNA-RT sont tous inclus dans la classe Revtraviricetes , phylum Arterviricota , royaume Pararnavirae du royaume Riboviria . À l'exception des Caulimoviridae , qui appartiennent au groupe VII, tous les membres de l' ordre Revtraviricetes Ortervirales sont des virus ssRNA-RT.
Groupe VII : virus à ADN double brin avec un intermédiaire ARN
Le septième groupe de Baltimore contient des virus qui ont un génome d'ADN double brin qui a un intermédiaire d'ARN (dsADN-RT) dans son cycle de réplication. Les virus dsADN-RT ont un trou dans un brin, qui est réparé pour créer un génome dsADN complet avant la transcription. Les virus dsADN-RT sont transcrits de la même manière que les virus dsADN, mais utilisent la transcription inverse pour répliquer leur génome circulaire alors qu'il est encore dans la capside. L'ARN polymérase II de la cellule hôte transcrit les brins d'ARN du génome dans le cytoplasme, et le génome est répliqué à partir de ces brins d'ARN. Le génome dsADN est produit à partir de brins d'ARN prégénomiques via le même mécanisme général que les virus ssARN-RT, mais avec une réplication se produisant dans une boucle autour du génome circulaire. Après la réplication, le génome dsADN peut être emballé ou envoyé au noyau pour d'autres cycles de transcription.
Les virus dsDNA-RT sont, comme les virus ssRNA-RT, tous inclus dans la classe Revtraviricetes . Deux familles de virus dsDNA-RT sont reconnues : Caulimoviridae , qui appartient à l'ordre des Ortervirales , et Hepadnaviridae , qui est la seule famille de l'ordre des Blubervirales .
Caractéristiques multi-groupes
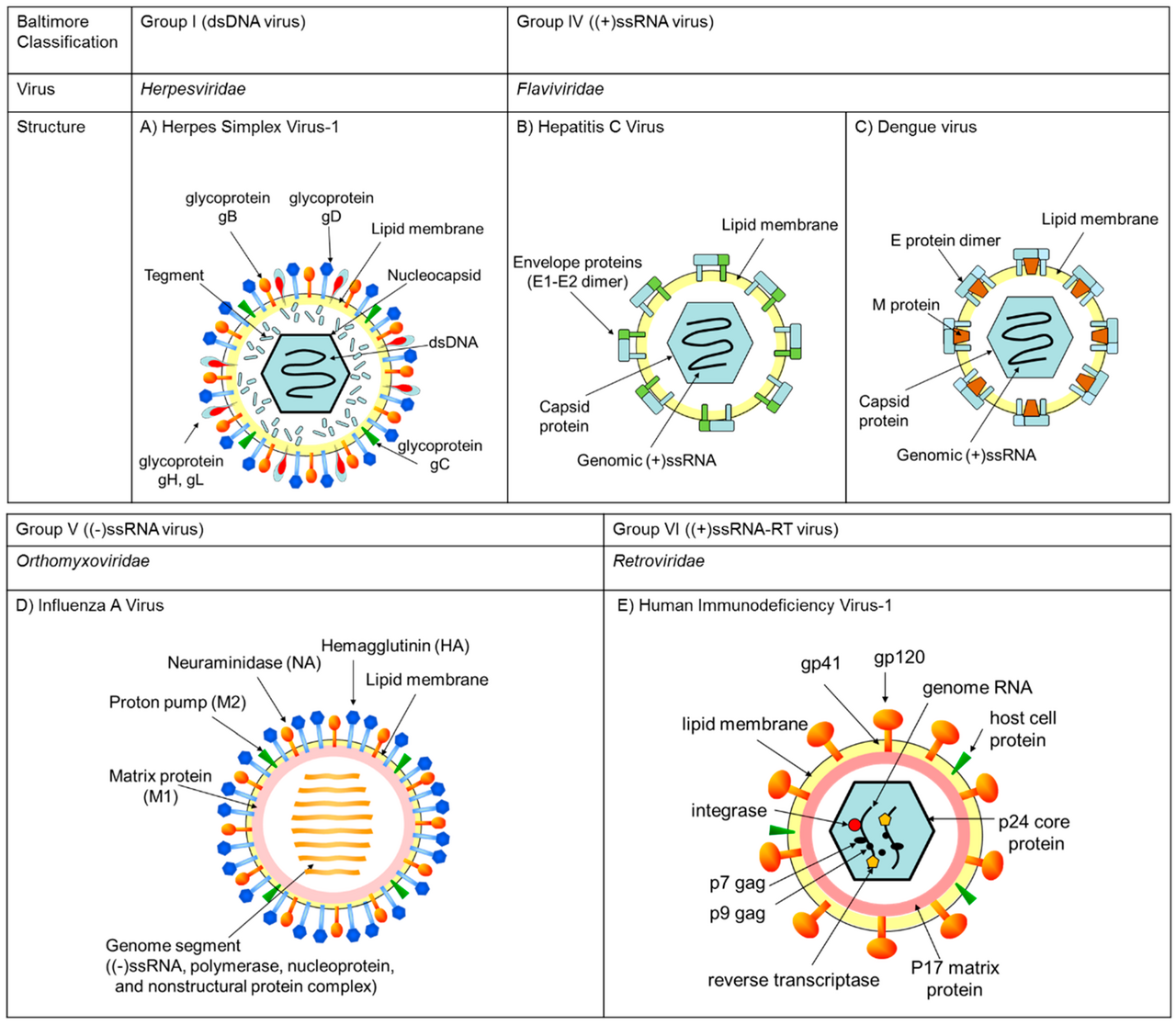
Un certain nombre de caractéristiques des virus ne sont pas directement associées à la classification de Baltimore, mais correspondent néanmoins étroitement à plusieurs groupes spécifiques de Baltimore. Cela comprend l'épissage alternatif pendant la transcription, la segmentation du génome viral, la gamme d'hôtes des virus, le caractère linéaire ou circulaire du génome et les différentes méthodes de traduction de l'ARNm viral.
Épissage alternatif
L'épissage alternatif est un mécanisme par lequel différentes protéines peuvent être produites à partir d'un seul gène en utilisant des sites d'épissage alternatifs pour produire différents ARNm. On le trouve dans divers virus à ADN, à ARN simple brin et à transcription inverse. Les virus peuvent utiliser l'épissage alternatif uniquement pour produire plusieurs protéines à partir d'un seul brin de pré-ARNm ou à d'autres fins spécifiques. Pour certains virus, notamment les familles Orthomyxoviridae et Papillomaviridae , l'épissage alternatif agit comme un moyen de réguler l'expression précoce et tardive des gènes au cours des différentes étapes de l'infection. Les herpèsvirus l'utilisent comme mécanisme de défense anti-hôte potentiel pour empêcher la synthèse de protéines antivirales spécifiques. De plus, en plus de l'épissage alternatif, comme l'ARN cellulaire non épissé ne peut pas être transporté hors du noyau, les hépadnavirus et les rétrovirus contiennent leurs propres protéines pour exporter leur ARN génomique non épissé hors du noyau.
Segmentation du génome
Les génomes viraux peuvent exister dans un seul segment, ou monopartite, ou ils peuvent être divisés en plusieurs molécules, appelées multipartites . Pour les virus monopartites, tous les gènes se trouvent sur le seul segment du génome. Les virus multipartites regroupent généralement leurs génomes dans un seul virion de sorte que l'ensemble du génome se trouve dans une seule particule virale et que les segments séparés contiennent des gènes différents. Les virus monopartites se trouvent dans tous les groupes de Baltimore, tandis que les virus multipartites sont généralement des virus à ARN. En effet, la plupart des virus multipartites infectent les plantes ou les champignons, qui sont des eucaryotes, et la plupart des virus eucaryotes sont des virus à ARN. La famille Pleolipoviridae varie car certains virus sont monopartites à ADN simple brin tandis que d'autres sont bipartites, un segment étant un ADN simple brin et l'autre un ADN double brin. Les virus de la famille des virus végétaux à ADN simple brin Geminiviridae varient également entre monopartites et bipartites.
Gamme d'hôtes
Différents groupes de Baltimore ont tendance à se retrouver dans différentes branches de la vie cellulaire. Chez les procaryotes, la grande majorité des virus sont des virus à ADN double brin, et une minorité significative sont des virus à ADN simple brin. Les virus à ARN procaryotes, en revanche, sont relativement rares. La plupart des virus eucaryotes, y compris la plupart des virus animaux et végétaux, sont des virus à ARN, bien que les virus à ADN eucaryotes soient également courants. Par groupe, la grande majorité des virus à ADN double brin infectent les procaryotes, les virus à ADN simple brin se trouvent dans les trois domaines de la vie, les virus à ARN double brin et à ARN simple brin + se trouvent principalement chez les eucaryotes mais aussi chez les bactéries, et les virus à ARN simple brin et à transcription inverse ne se trouvent que chez les eucaryotes.
Génomes linéaires et circulaires
Les génomes viraux peuvent être linéaires avec des extrémités ou circulaires en boucle. Le fait qu'un virus ait un génome linéaire ou circulaire varie d'un groupe à l'autre. Un pourcentage significatif de virus à ADN double brin sont les deux, les virus à ADN simple brin sont principalement circulaires, les virus à ARN et les virus à ARN simple brin-RT sont généralement linéaires, et les virus à ADN simple brin-RT sont généralement circulaires. des Sphaerolipoviridae à ADN double brin et dans la famille des Pleolipoviridae , les virus contiennent à la fois des génomes linéaires et circulaires, variant d'un genre à l'autre.
Édition d'ARN
L'édition de l'ARN est utilisée par divers virus à ARN simple brin pour produire différentes protéines à partir d'un seul gène. Cela peut être réalisé par glissement de la polymérase pendant la transcription ou par édition post-transcriptionnelle. Dans le cas du glissement de la polymérase, l'ARN polymérase glisse d'un nucléotide en arrière pendant la transcription, insérant un nucléotide non inclus dans le brin modèle. L'édition d'un modèle génomique altérerait l'expression des gènes, de sorte que l'édition de l'ARN n'est effectuée que pendant et après la transcription. Pour les virus Ebola , l'édition de l'ARN améliore la capacité d'adaptation à leurs hôtes.
L'épissage alternatif diffère de l'édition d'ARN dans la mesure où l'épissage alternatif ne modifie pas la séquence d'ARNm comme l'édition d'ARN, mais modifie plutôt la capacité de codage d'une séquence d'ARNm en raison de sites d'épissage alternatifs. Les deux mécanismes ont par ailleurs le même résultat : plusieurs protéines sont exprimées à partir d'un seul gène.
Traduction
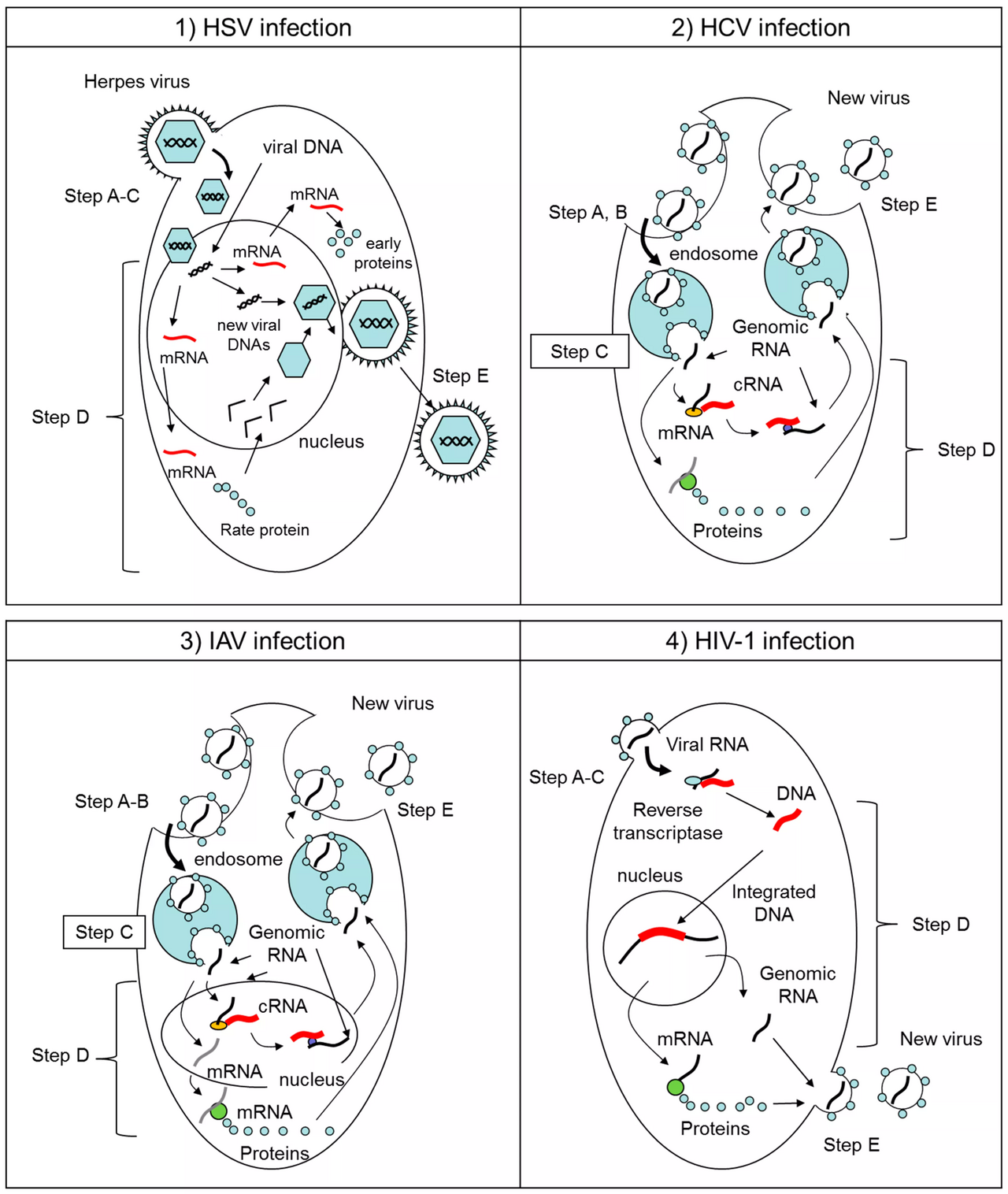
La traduction est le processus par lequel les protéines sont synthétisées à partir de l'ARNm par les ribosomes . Les groupes Baltimore ne se rapportent pas directement à la traduction des protéines virales, mais divers types atypiques de traduction utilisés par les virus se trouvent généralement dans des groupes Baltimore spécifiques :
- Initiation de la traduction non canonique :
- Initiation virale de la traduction : utilisée principalement par les virus +ssRNA et ssRNA-RT, divers virus ont développé des mécanismes pour initier la traduction, tels que des sites d'entrée ribosomiques internes pour permettre une traduction indépendante de la coiffe, des boucles en épingle à cheveux en aval qui permettent une traduction dépendante de la coiffe en l'absence d'un facteur d'initiation eIF2 , et l'initiation au niveau d'un CUG ou d'un autre codon de départ avec un acide aminé leucine .
- Balayage par fuite : utilisé par divers virus dans tous les groupes Baltimore, la sous-unité ribosomique 40S peut balayer un codon de départ, sautant ainsi un ORF, initiant uniquement la traduction avec la sous-unité 60S à un codon de départ ultérieur.
- Dérivation ribosomique : utilisée par divers virus dsADN, +ssARN, -ssARN, ssARN-RT, a dsADN-RT, les ribosomes commenceront à scanner à partir d'une structure de coiffe 5' puis contourneront une structure leader dans l'ARNm, initiant la traduction en aval de la séquence leader.
- Terminaison-réinitiation : utilisé par certains virus à dsRNA et +ssRNA, les ribosomes peuvent traduire un ORF, mais après la terminaison de la traduction de cet ORF, une proportion de sous-unités 40S du ribosome reste attachée à l'ARNm comme moyen de relancer la traduction d'un ORF ultérieur.
- Allongement non canonique et terminaison de la traduction :
- Décalage du cadre ribosomique : utilisé par divers virus à ADN double brin, à ARN double brin, à ARN simple brin et à ARN simple brin-RT, il produit des protéines fusionnées à partir d'ORFs superposés. Cela est exécuté simplement par les ribosomes qui glissent une nucléobase vers l'avant ou vers l'arrière pendant la traduction.
- Suppression de la terminaison : également appelée lecture continue du codon stop, utilisée par divers virus dsRNA, +ssRNA et ssRNA-RT, certains virus contiennent des codons dans leur ARNm qui signaleraient normalement la fin de la traduction lorsqu'ils sont reconnus par un facteur de libération , mais qui sont en fait partiellement reconnus par l'ARNt pendant la traduction, ce qui permet une traduction continue jusqu'au prochain codon stop afin de produire une extrémité étendue de la protéine virale. Chez les virus, cela est souvent utilisé pour exprimer des enzymes de réplication .
- Ribosomal skipping : également appelé stop-carry on, utilisé par divers virus à ARNdb et à ARNss+, un peptide viral ou une séquence d'acides aminés peut empêcher un ribosome de se lier de manière covalente à un nouvel acide aminé inséré, ce qui bloque la traduction ultérieure. Par conséquent, la polyprotéine est clivée de manière co-traductionnelle et une nouvelle séquence d'acides aminés est démarrée, ce qui conduit à la production de deux protéines individuelles à partir d'un ORF.
Histoire

La classification de Baltimore a été proposée en 1971 par le virologue David Baltimore dans un article intitulé Expression of Animal Virus Genomes . Elle contenait initialement les six premiers groupes, mais a ensuite été élargie pour inclure le groupe VII. En raison de l'utilité de la classification de Baltimore, elle a été utilisée parallèlement à la taxonomie virale standard, qui est basée sur les relations évolutives et régie par le Comité international de taxonomie des virus (ICTV).
Entre les années 1990 et 2010, la taxonomie des virus utilisait un système à 5 rangs allant de l'ordre à l'espèce, avec la classification de Baltimore utilisée conjointement. En dehors du cadre officiel de l'ICTV, divers supergroupes de virus rejoignant différentes familles et ordres ont été créés au fil du temps sur la base de preuves croissantes de relations évolutives plus profondes. Par conséquent, en 2016, l'ICTV a commencé à envisager d'établir des rangs supérieurs à l'ordre ainsi que la manière dont les groupes de Baltimore seraient traités parmi les taxons supérieurs.
Lors de deux votes en 2018 et 2019, un système de 15 rangs allant du royaume à l'espèce a été établi par l'ICTV. Dans ce cadre, les groupes Baltimore pour les virus à ARN et les virus RT ont été intégrés dans des taxons formels. En 2018, le royaume Riboviria a été établi et comprenait initialement les trois groupes de virus à ARN. Un an plus tard, Riboviria a été élargi pour inclure également les deux groupes RT. Au sein du royaume, les virus RT sont inclus dans le royaume Pararnavirae et les virus à ARN dans le royaume Orthornavirae . De plus, les trois groupes Baltimore pour les virus à ARN sont utilisés comme caractéristiques définissant les phylums dans Orthornavirae .
Contrairement aux virus à ARN et aux virus RT, les virus à ADN ne sont pas réunis sous un seul règne mais sont plutôt dispersés dans quatre règnes et divers taxons qui ne sont pas assignés à un règne. Les règnes Adnaviria et Duplodnaviria contiennent exclusivement des virus à ADN double brin, Monodnaviria contient principalement des virus à ADN simple brin, mais contient également des virus à ADN simple brin, et Varidnaviria contient exclusivement des virus à ADN simple brin, bien que certains membres proposés de Varidnaviria , à savoir la famille Finnlakeviridae , soient des virus à ADN simple brin.