cellule à se différencier en d'autres types cellulaires . Plus une cellule peut se différencier en différents types cellulaires, plus sa potentialité est élevée. La potentialité est également décrite comme le potentiel d'activation des gènes au sein d'une cellule, qui, tel un continuum, commence par la totipotence pour désigner une cellule possédant le plus grand potentiel de différenciation, puis la pluripotence , la multipotence , l'oligopotence et enfin l'unipotence .
Une cellule pleinement différenciée peut retrouver un état de totipotence. La conversion vers la totipotence est complexe et encore mal comprise. En 2011, des recherches ont révélé que les cellules peuvent se différencier non pas en une cellule pleinement totipotente, mais plutôt en une « variation cellulaire complexe » de la totipotence.
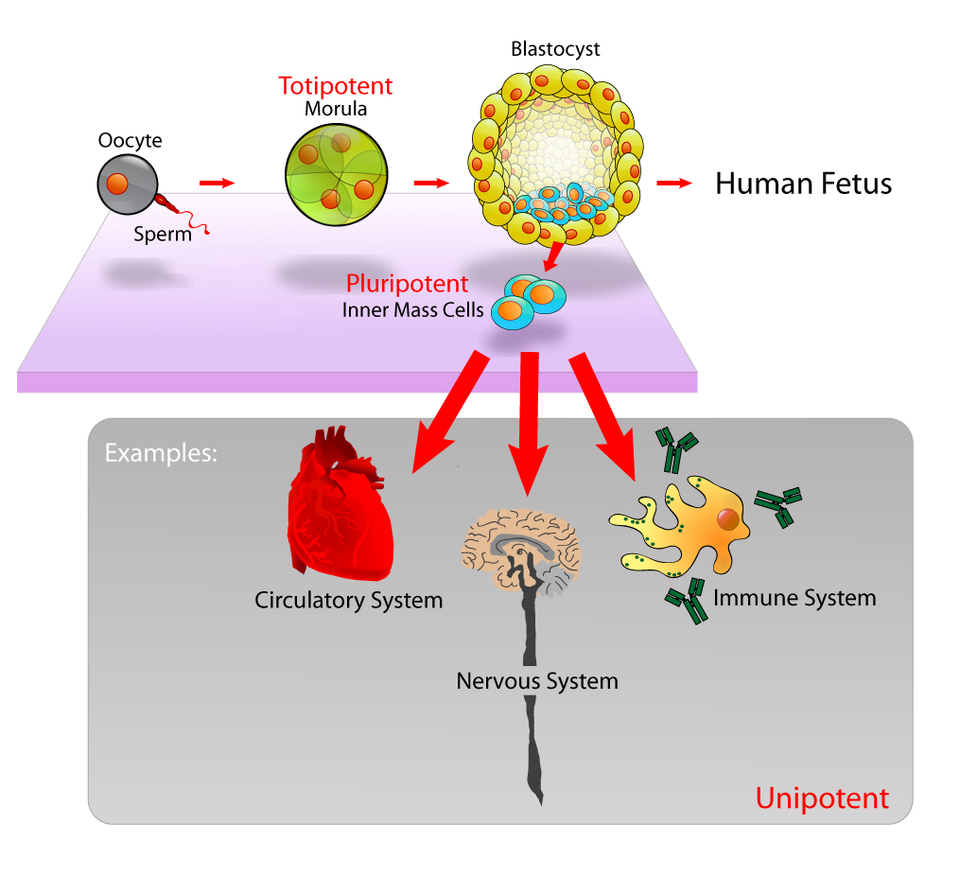
Le modèle du développement humain permet de décrire l'origine des cellules totipotentes. Le développement humain débute lorsqu'un spermatozoïde féconde un ovule. L'ovule fécondé ainsi formé donne naissance à une unique cellule totipotente, le zygote . Dans les premières heures suivant la fécondation, ce zygote se divise en cellules totipotentes identiques, qui peuvent ensuite se différencier en l'un des trois feuillets embryonnaires humains ( endoderme , mésoderme ou ectoderme ), ou en cellules du placenta ( cytotrophoblaste ou syncytiotrophoblaste ). Après avoir atteint le stade de 16 cellules, les cellules totipotentes de la morula se différencient en cellules qui deviendront soit la masse cellulaire interne du blastocyste , soit les trophoblastes externes . Environ quatre jours après la fécondation, et après plusieurs cycles de division cellulaire, ces cellules totipotentes commencent à se spécialiser. La masse cellulaire interne, source des cellules souches embryonnaires , acquiert la pluripotence.
Des recherches sur Caenorhabditis elegans suggèrent que de multiples mécanismes, dont la régulation de l'ARN , pourraient jouer un rôle dans le maintien de la totipotence à différents stades de développement chez certaines espèces. Des travaux menés sur le poisson-zèbre et les mammifères suggèrent une interaction supplémentaire entre les microARN et les protéines de liaison à l'ARN (RBP) dans la détermination des différences de développement.
Cellules germinales primordiales
Chez la souris , la reprogrammation génomique à l'échelle du génome , conduisant à la totipotence, implique l'effacement des empreintes épigénétiques . Cette reprogrammation est facilitée par une déméthylation active de l'ADN via la voie enzymatique de réparation par excision de base de l'ADN . Cette voie implique l'effacement de la méthylation des îlots CpG (5mC) dans les cellules germinales primordiales par la conversion initiale de la 5mC en 5-hydroxyméthylcytosine (5hmC), une réaction catalysée par les enzymes ten-eleven dioxygénases TET-1 et TET-2 , présentes en grande quantité
Pluripotence
Une cellule souche pluripotente ( cellule souche qui a le potentiel de se différencier en n'importe laquelle des cellules des trois feuillets germinatifs : endoderme (intestin, poumons et foie), mésoderme (muscle, squelette, système vasculaire sanguin, système urogénital, derme) ou ectoderme (système nerveux, système sensoriel, épiderme), mais pas en tissus extra-embryonnaires comme le placenta ou le sac vitellin.
pluripotence induite
On pense également que des facteurs épigénétiques interviennent dans la reprogrammation des cellules somatiques en vue d'induire la pluripotence. Selon l'hypothèse, certains facteurs épigénétiques pourraient effacer les marques épigénétiques somatiques initiales afin d'acquérir les nouvelles marques nécessaires à l'obtention d'un état pluripotent. La chromatine est également réorganisée dans les cellules iPS et devient similaire à celle des cellules ESC : moins condensée, elle est donc plus accessible. Les modifications de l'euchromatine sont également fréquentes, ce qui concorde avec l'état de l'euchromatine observé dans les cellules ESC.
En raison de leur grande similarité avec les cellules souches embryonnaires (CSE), les communautés médicales et de recherche s'intéressent aux cellules souches pluripotentes induites (iPSC). Les iPSC pourraient potentiellement avoir les mêmes implications et applications thérapeutiques que les CSE, mais sans l'utilisation controversée d'embryons, un sujet de vifs débats bioéthiques. La pluripotence induite de cellules somatiques en cellules iPS indifférenciées a initialement été saluée comme la fin de l' utilisation controversée des cellules souches embryonnaires . Cependant, les iPSC se sont révélées potentiellement tumorigènes et, malgré des progrès , n'ont jamais été approuvées pour la recherche clinique aux États-Unis jusqu'à récemment. Actuellement, des cellules progénitrices dopaminergiques autologues dérivées d'iPSC sont utilisées dans des essais cliniques pour le traitement de la maladie de Parkinson . Des obstacles tels que de faibles taux de réplication et une sénescence précoce ont également été rencontrés lors de la production d'iPSC , limitant leur utilisation comme substituts aux CSE.
L'expression somatique de facteurs de transcription combinés peut induire directement d'autres destins cellulaires somatiques définis ( transdifférenciation ) ; des chercheurs ont identifié trois facteurs de transcription spécifiques de la lignée neuronale capables de convertir directement des fibroblastes de souris (cellules du tissu conjonctif) en neurones pleinement fonctionnels . Ce résultat remet en question la nature terminale de la différenciation cellulaire et l'intégrité de l'engagement de lignage ; et suggère qu'avec les outils appropriés, toutes les cellules sont totipotentes et peuvent former tous types de tissus.
Parmi les applications médicales et thérapeutiques potentielles des cellules souches pluripotentes induites (iPSC) dérivées de patients, on peut citer leur utilisation dans les transplantations cellulaires et tissulaires, sans le risque de rejet fréquemment rencontré. Les iPSC pourraient potentiellement remplacer les modèles animaux inadaptés ainsi que les modèles in vitro utilisés pour la recherche sur les maladies.
Essais de formation de tératomes

Avec l'essor des recherches et des applications des cellules souches embryonnaires (ESC) et des cellules souches pluripotentes induites (iPSC) en médecine régénérative, le contrôle qualité des cellules testées est indispensable. Le test de formation de tératomes est une procédure largement acceptée, applicable aussi bien aux ESC de mammifères qu'aux iPSC. Un tératome est une tumeur généralement bénigne caractérisée par sa capacité à former les trois feuillets embryonnaires : l'ectoderme (nerfs, épithélium), le mésoderme (muscles, os et cartilage) et l'endoderme (intestin).
Un test de formation de tératomes consiste à injecter des cellules souches pluripotentes dans différents tissus. Parmi ces tissus, on peut citer la capsule rénale, les régions intratesticulaires et intramusculaires de souris immunodéficientes. La pluripotence est caractérisée par la capacité des cellules souches à former un tératome capable de produire les trois feuillets embryonnaires.
Bien que le test de formation de tératomes soit considéré comme la méthode de référence par les chercheurs, de nombreuses difficultés ont été rencontrées. L'une d'elles réside dans le manque de standardisation des détails et des facteurs influençant la formation de tératomes. Parmi les points à standardiser figurent les sites de greffe , l'âge de l'organisme testé (généralement des souris) et le nombre de cellules injectées. Ces tests sont également coûteux et complexes à mettre en œuvre, et l'utilisation d'organismes tests soulève des questions éthiques.
Un autre problème lié à ce type de test est le risque d'erreurs d'interprétation histologique. Des cellules qui ne sont pas complètement reprogrammées en cellules iPS formant des amas cellulaires visibles, présentant des caractéristiques similaires à celles des tératomes, peuvent être considérées comme pluripotentes alors qu'elles sont dépourvues des trois feuillets embryonnaires. La nécessité de suivre les lignées cellulaires et de distinguer les cellules hôtes des cellules donneuses a également été soulignée. Certains réactifs de préparation cellulaire peuvent induire une réponse inflammatoire ou une réponse immunitaire à un antigène étranger . Ces réponses peuvent contribuer à une identification erronée de la différenciation des cellules testées.
États de pluripotence naïfs vs. amorcés
Les résultats concernant les épiblastes avant et après implantation ont conduit à proposer une classification de la pluripotence en deux états : « naïf » et « primé », correspondant respectivement à l’épiblaste pré- et post-implantation. Le passage de l’état naïf à l’état primé est contrôlé par la réduction de la dimérisation Sox2/Oct4 sur les éléments d’ADN SoxOct qui régulent la pluripotence naïve. Des cellules souches pluripotentes primées, issues de différentes espèces, peuvent être ramenées à l’état naïf grâce à un cocktail contenant Klf4 et Sox2, ou « super-Sox » – un facteur de transcription chimérique présentant une capacité accrue de dimérisation avec Oct4.
Les cellules souches de base couramment utilisées en science, appelées cellules souches embryonnaires (CSE), dérivent de l'épiblaste pré-implantatoire. Cet épiblaste est capable de générer le fœtus entier, et une seule cellule épiblastique peut contribuer à toutes les lignées cellulaires si elle est injectée dans un autre blastocyste. Par ailleurs, plusieurs différences notables existent entre les épiblastes pré- et post-implantatoires, notamment au niveau de leur morphologie. Après l'implantation, l'épiblaste adopte une forme de coupe appelée « cylindre ovocytaire », et au niveau des chromosomes, avec l'inactivation aléatoire de l'un des chromosomes X au début du développement du cylindre ovocytaire ( inactivation du chromosome X) . Au cours de ce développement, les cellules épiblastiques du cylindre ovocytaire sont systématiquement ciblées par les facteurs de croissance des fibroblastes , la voie de signalisation Wnt et d'autres facteurs inducteurs via le sac vitellin et le tissu trophoblastique environnants , ce qui leur confère une spécificité instructive en fonction de leur organisation spatiale.
Une autre différence majeure réside dans l'incapacité des cellules souches de l'épiblaste post-implantation à contribuer à la formation de chimères blastocystiques , ce qui les distingue des autres cellules souches pluripotentes connues. Les lignées cellulaires dérivées de ces épiblastes post-implantation sont appelées cellules souches dérivées de l'épiblaste (EpiSC) , obtenues pour la première fois en laboratoire en 2007. Les cellules souches embryonnaires (ESC) et les cellules souches épiblastiques (EpiSC) dérivent toutes deux de l'épiblaste, mais à des stades de développement différents. La pluripotence est encore intacte dans l'épiblaste post-implantation, comme le démontre l'expression conservée de Nanog , Oct-4 dans les EpiSC , jusqu'à la somitogenèse , et peut être inversée en cours d'expression induite d' Oct-4 .
pluripotence native chez les plantes

Une pluripotence non induite a été observée dans des cultures de tissus PLETHORA1 et PLETHORA2 , ainsi que PLETHORA3 , PLETHORA5 et PLETHORA7 , dont l'expression a été induite par l'auxine , selon Kareem . (Ces régulateurs sont également connus sous les noms de PLT1, PLT2, PLT3, PLT5 et PLT7, et sont codés par des gènes portant les mêmes noms.) ( La multipotence désigne la capacité des cellules progénitrices à activer leurs gènes pour se différencier en différents types cellulaires. Par exemple, une cellule souche hématopoïétique peut se différencier en plusieurs types de cellules sanguines, comme les lymphocytes , les monocytes , les neutrophiles , etc. Cependant, on ignore encore si les cellules souches hématopoïétiques peuvent se différencier en cellules cérébrales , en cellules osseuses ou en d'autres types cellulaires non sanguins.le sang de cordon , le tissu adipeux , les cellules cardiaques , la moelle osseuse et les cellules souches mésenchymateuses (CSM) qui se trouvent dans la troisième molaire . Les CSM pourraient constituer une source précieuse de cellules souches à partir des molaires chez les enfants de 8 à 10 ans, avant la calcification dentaire adulte. Les CSM peuvent se différencier en ostéoblastes, chondrocytes et adipocytes.
oligopotence
En biologie, l'oligopotence est la capacité des cellules progénitrices à se différencier en quelques types cellulaires . Il s'agit d'un degré de potentialité . Les cellules souches lymphoïdes et myéloïdes sont des exemples de cellules souches oligopotentes. Une cellule lymphoïde, en particulier, peut donner naissance à diverses cellules sanguines, telles que les lymphocytes B et T, mais pas à un autre type de cellule sanguine comme un globule rouge. Les cellules souches vasculaires, capables de se différencier en cellules endothéliales ou en cellules musculaires lisses, sont des exemples de cellules progénitrices.
Unipotence
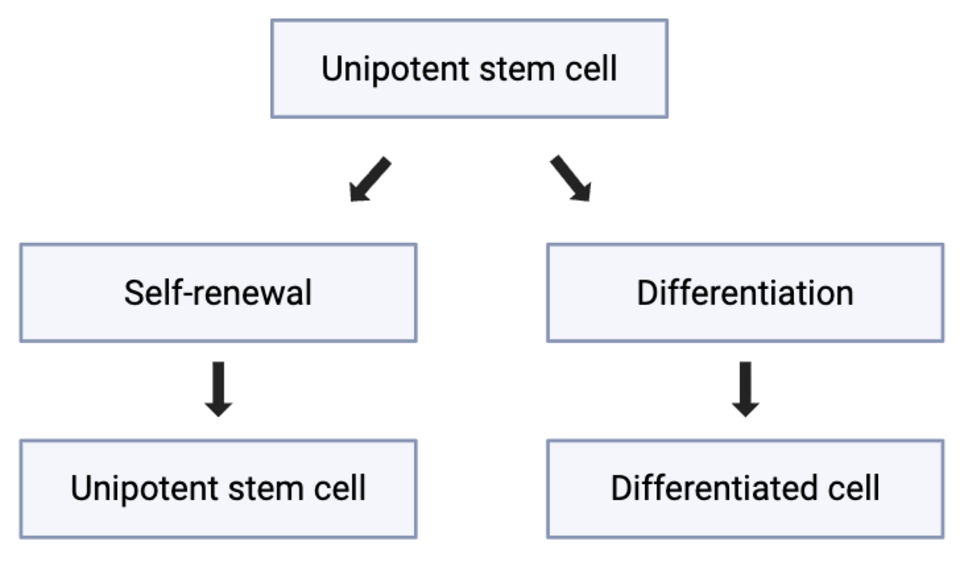
Cependant, la classification des cellules souches véritablement unipotentes demeure un sujet de débat scientifique. Certaines cellules précédemment qualifiées d'unipotentes pourraient en réalité être des cellules progénitrices . Les cellules progénitrices sont des cellules déjà engagées dans une voie de développement spécifique et dépourvues de capacité d'auto-renouvellement à long terme . Cette distinction est importante car les véritables cellules souches doivent être capables à la fois de se différencier et de maintenir leur population au fil du temps . Par conséquent, l'identification d'une cellule véritablement unipotente peut s'avérer complexe.
Par exemple, on pensait autrefois que les hépatoblastes étaient unipotents. En réalité, ils sont bipotents car ils peuvent se différencier en hépatocytes ou en cholangiocytes . Cet exemple illustre comment les progrès des données expérimentales peuvent affiner la classification des types cellulaires.
nullipotence
En biologie cellulaire, une cellule nullipotente est une cellule incapable de se différencier en un autre type cellulaire ( les neurones , les globules rouges , etc.), il est le plus souvent employé pour désigner un carcinome embryonnaire (CE) ou des cellules souches embryonnaires ayant perdu leur capacité de différenciation (généralement suite à des mutations génétiques)