
Un vecteur de clonage est un petit fragment d' ADN qui peut être maintenu de manière stable dans un organisme et dans lequel un fragment d'ADN étranger peut être inséré à des fins de clonage . Le vecteur de clonage peut être de l'ADN provenant d'un virus , d'une cellule d'un organisme supérieur ou du plasmide d'une bactérie. Le vecteur contient des caractéristiques qui permettent l'insertion pratique d'un fragment d'ADN dans le vecteur ou son retrait du vecteur, par exemple grâce à la présence de sites de restriction . Le vecteur et l'ADN étranger peuvent être traités avec une enzyme de restriction qui coupe l'ADN et les fragments d'ADN ainsi générés contiennent soit des extrémités franches, soit des surplombs appelés extrémités collantes, et l'ADN vecteur et l'ADN étranger ayant des extrémités compatibles peuvent ensuite être joints par ligature moléculaire . Une fois qu'un fragment d'ADN a été cloné dans un vecteur de clonage, il peut être sous-cloné dans un autre vecteur conçu pour une utilisation plus spécifique.
Il existe de nombreux types de vecteurs de clonage, mais les plus couramment utilisés sont les plasmides génétiquement modifiés . Le clonage est généralement réalisé en premier lieu à l'aide d'Escherichia coli , et les vecteurs de clonage dans E. coli comprennent les plasmides, les bactériophages (tels que le phage λ ), les cosmides et les chromosomes artificiels bactériens (BAC). Cependant, certains ADN ne peuvent pas être maintenus de manière stable dans E. coli , par exemple les très gros fragments d'ADN, et d'autres organismes tels que la levure peuvent être utilisés. Les vecteurs de clonage dans la levure comprennent les chromosomes artificiels de levure (YAC).
Caractéristiques d'un vecteur de clonage
Tous les vecteurs de clonage couramment utilisés en biologie moléculaire possèdent des caractéristiques clés nécessaires à leur fonction, comme un site de clonage adapté et un marqueur sélectionnable. D'autres peuvent avoir des caractéristiques supplémentaires spécifiques à leur utilisation. Pour des raisons de facilité et de commodité, le clonage est souvent réalisé en utilisant E. coli . Ainsi, les vecteurs de clonage utilisés possèdent souvent des éléments nécessaires à leur propagation et à leur maintien dans E. coli , comme une origine de réplication fonctionnelle (ori). L' origine de réplication ColE1 se trouve dans de nombreux plasmides. Certains vecteurs comprennent également des éléments qui leur permettent d'être maintenus dans un autre organisme en plus d' E. coli , et ces vecteurs sont appelés vecteur navette .
Site de clonage
Tous les vecteurs de clonage ont des caractéristiques qui permettent d'insérer facilement un gène dans le vecteur ou de le retirer de celui-ci. Il peut s'agir d'un site de clonage multiple (MCS) ou d'un polylinker, qui contient de nombreux sites de restriction uniques . Les sites de restriction dans le MCS sont d'abord clivés par des enzymes de restriction, puis un gène cible amplifié par PCR également digéré avec les mêmes enzymes est ligaturé dans les vecteurs à l'aide d' une ligase d'ADN . La séquence d'ADN cible peut être insérée dans le vecteur dans une direction spécifique si cela est souhaité. Les sites de restriction peuvent être utilisés ultérieurement pour le sous-clonage dans un autre vecteur si nécessaire.
D'autres vecteurs de clonage peuvent utiliser la topoisomérase au lieu de la ligase et le clonage peut être réalisé plus rapidement sans nécessiter de digestion de restriction du vecteur ou d'insertion. Dans cette méthode de clonage TOPO, un vecteur linéarisé est activé en attachant la topoisomérase I à ses extrémités, et ce vecteur « activé par TOPO » peut ensuite accepter un produit PCR en ligaturant les deux extrémités 5' du produit PCR, libérant la topoisomérase et formant un vecteur circulaire dans le processus. Une autre méthode de clonage sans utilisation de digestion d'ADN et de ligase est la recombinaison d'ADN , par exemple comme utilisée dans le système de clonage Gateway . Le gène, une fois cloné dans le vecteur de clonage (appelé clone d'entrée dans cette méthode), peut être introduit de manière pratique dans une variété de vecteurs d'expression par recombinaison.
Marqueur sélectionnable
Un marqueur sélectionnable est porté par le vecteur pour permettre la sélection des cellules transformées positivement . La résistance aux antibiotiques est souvent utilisée comme marqueur, par exemple le gène de la bêta-lactamase , qui confère une résistance au groupe de pénicilline des antibiotiques bêta-lactamines comme l'ampicilline . Certains vecteurs contiennent deux marqueurs sélectionnables, par exemple le plasmide pACYC177 possède à la fois un gène de résistance à l'ampicilline et à la kanamycine . Le vecteur navette qui est conçu pour être maintenu dans deux organismes différents peut également nécessiter deux marqueurs sélectionnables, bien que certains marqueurs sélectionnables tels que la résistance à la zéocine et à l'hygromycine B soient efficaces dans différents types de cellules. Des marqueurs de sélection auxotrophiques qui permettent à un organisme auxotrophe de se développer dans un milieu de croissance minimal peuvent également être utilisés ; des exemples de ceux-ci sont LEU2 et URA3 qui sont utilisés avec leurs souches auxotrophes de levure correspondantes.
Un autre type de marqueur sélectionnable permet la sélection positive du plasmide avec le gène cloné. Cela peut impliquer l'utilisation d'un gène mortel pour les cellules hôtes, comme la barnase , Ccda , et les toxines parD/parE . Cela fonctionne généralement en perturbant ou en supprimant le gène mortel pendant le processus de clonage, et les clones non réussis où le gène mortel reste intact tueraient les cellules hôtes, donc seuls les clones réussis sont sélectionnés.
Gène rapporteur
Les gènes rapporteurs sont utilisés dans certains vecteurs de clonage pour faciliter le criblage des clones réussis en utilisant les caractéristiques de ces gènes qui permettent d'identifier facilement les clones réussis. De telles caractéristiques présentes dans les vecteurs de clonage peuvent être le fragment lacZ α pour la complémentation α dans la sélection bleu-blanc , et/ou le gène marqueur ou les gènes rapporteurs dans le cadre et encadrant le MCS pour faciliter la production de protéines de fusion . Des exemples de partenaires de fusion qui peuvent être utilisés pour le criblage sont la protéine fluorescente verte (GFP) et la luciférase .
Éléments d'expression
Un vecteur de clonage ne doit pas nécessairement contenir des éléments appropriés pour l' expression d'un gène cible cloné, tels qu'un promoteur et un site de liaison ribosomique (RBS), mais beaucoup le font et peuvent alors fonctionner comme un vecteur d'expression . L' ADN cible peut être inséré dans un site qui est sous le contrôle d'un promoteur particulier nécessaire à l'expression du gène cible dans l'hôte choisi. Lorsque le promoteur est présent, l'expression du gène est de préférence étroitement contrôlée et inductible de sorte que les protéines ne sont produites que lorsque cela est nécessaire. Certains promoteurs couramment utilisés sont les promoteurs T7 et lac . La présence d'un promoteur est nécessaire lorsque des techniques de criblage telles que la sélection bleu-blanc sont utilisées.
Des vecteurs de clonage sans promoteur ni RBS pour la séquence d'ADN clonée sont parfois utilisés, par exemple lors du clonage de gènes dont les produits sont toxiques pour les cellules d'E. coli . Le promoteur et le RBS pour la séquence d'ADN clonée sont également inutiles lors de la première création d'une bibliothèque génomique ou d'ADNc de clones, car les gènes clonés sont normalement sous-clonés dans un vecteur d'expression plus approprié si leur expression est requise.
Certains vecteurs sont conçus pour la transcription uniquement, sans expression de protéine hétérologue, par exemple pour la production d'ARNm in vitro . Ces vecteurs sont appelés vecteurs de transcription. Ils peuvent ne pas contenir les séquences nécessaires à la polyadénylation et à la terminaison, et ne peuvent donc pas être utilisés pour la production de protéines.
Types de vecteurs de clonage
Il existe un grand nombre de vecteurs de clonage, et le choix du vecteur peut dépendre d'un certain nombre de facteurs, tels que la taille de l'insert, le nombre de copies et la méthode de clonage. Un insert de grande taille peut ne pas être maintenu de manière stable dans un vecteur de clonage général, en particulier pour ceux qui ont un nombre de copies élevé, c'est pourquoi le clonage de grands fragments peut nécessiter un vecteur de clonage plus spécialisé.
Plasmide
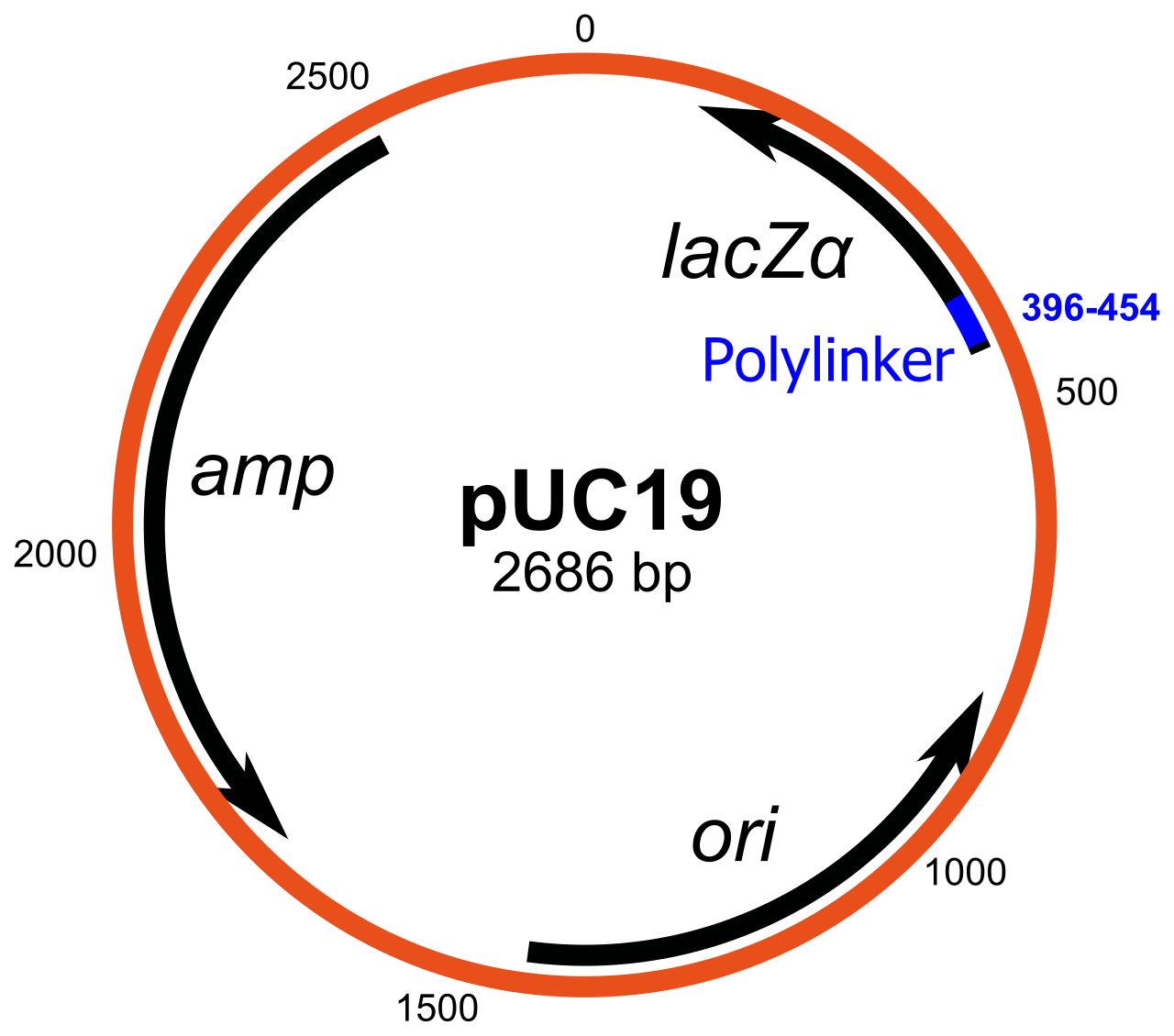
Les plasmides sont des ADN extrachromosomiques circulaires à réplication autonome. Ce sont les vecteurs de clonage standard et les plus couramment utilisés. La plupart des plasmides généraux peuvent être utilisés pour cloner des inserts d'ADN d'une taille allant jusqu'à 15 kb. L'un des premiers vecteurs de clonage couramment utilisés est le plasmide pBR322 . D'autres vecteurs de clonage incluent la série de plasmides pUC , et un grand nombre de vecteurs de plasmides de clonage différents sont disponibles. De nombreux plasmides ont un nombre de copies élevé, par exemple, pUC19 a un nombre de copies de 500 à 700 copies par cellule, et un nombre de copies élevé est utile car il produit un rendement plus élevé de plasmide recombinant pour une manipulation ultérieure. Cependant, des plasmides à faible nombre de copies peuvent être utilisés de préférence dans certaines circonstances, par exemple, lorsque la protéine du gène cloné est toxique pour les cellules.
Certains plasmides contiennent une origine de réplication du bactériophage M13 et peuvent être utilisés pour générer de l'ADN simple brin. On les appelle phagemides , et la série de vecteurs de clonage pBluescript en est un exemple.
Bactériophage
Les bactériophages utilisés pour le clonage sont le phage λ et le phage M13 . Il existe une limite supérieure à la quantité d'ADN qui peut être emballée dans un phage (un maximum de 53 kb), donc pour permettre à l'ADN étranger d'être inséré dans l'ADN du phage, les vecteurs de clonage de phage peuvent avoir besoin de supprimer certains gènes non essentiels, par exemple les gènes de la lysogénie puisque l'utilisation du phage λ comme vecteur de clonage implique uniquement le cycle lytique. Il existe deux types de vecteurs de phage λ - vecteur d'insertion et vecteur de remplacement. Les vecteurs d'insertion contiennent un site de clivage unique par lequel un ADN étranger d'une taille de 5 à 11 kb peut être inséré. Dans les vecteurs de remplacement, les sites de clivage flanquent une région contenant des gènes non essentiels au cycle lytique, et cette région peut être supprimée et remplacée par l'insert d'ADN dans le processus de clonage, et un ADN de plus grande taille de 8 à 24 kb peut être inséré.
Il existe également une limite inférieure de taille pour l'ADN qui peut être emballé dans un phage, et l'ADN vecteur qui est trop petit ne peut pas être correctement emballé dans le phage. Cette propriété peut être utilisée pour la sélection - un vecteur sans insert peut être trop petit, donc seuls les vecteurs avec insert peuvent être sélectionnés pour la propagation.
Cosmide
Les cosmides sont des plasmides qui incorporent un segment d'ADN du bactériophage λ qui possède le site terminal cohésif ( cos ) qui contient les éléments nécessaires à l'emballage de l'ADN dans les particules λ. Sous une origine de réplication appropriée (ori), il peut se répliquer sous forme de plasmide. Il est normalement utilisé pour cloner de grands fragments d'ADN entre 28 et 45 Kb.
Chromosome artificiel bactérien
Des tailles d'insertion allant jusqu'à 350 kb peuvent être clonées dans un chromosome artificiel bactérien (BAC). Les BAC sont conservés dans E. coli avec un nombre de copies de seulement 1 par cellule. Les BAC sont basés sur le plasmide F , un autre chromosome artificiel appelé PAC est basé sur le phage P1 .
Chromosome artificiel de levure
Les chromosomes artificiels de levure sont utilisés comme vecteurs pour cloner des fragments d'ADN de plus d'une mégabase (1 Mo = 1 000 kb). Ils sont utiles pour cloner des fragments d'ADN plus grands, comme l'exige la cartographie des génomes, comme dans le projet du génome humain . Ils contiennent une séquence télomérique, une séquence à réplication autonome (caractéristiques requises pour répliquer des chromosomes linéaires dans les cellules de levure). Ces vecteurs contiennent également des sites de restriction appropriés pour cloner de l'ADN étranger ainsi que des gènes à utiliser comme marqueurs sélectionnables.
Chromosome artificiel humain
Le chromosome artificiel humain peut être potentiellement utile comme vecteur de transfert de gènes pour l'introduction de gènes dans des cellules humaines, et comme outil pour les études d'expression et la détermination de la fonction des chromosomes humains. Il peut transporter de très gros fragments d'ADN (il n'y a pas de limite supérieure de taille à des fins pratiques), il ne présente donc pas le problème de la capacité de clonage limitée d'autres vecteurs, et il évite également une éventuelle mutagenèse insertionnelle causée par l'intégration dans les chromosomes de l'hôte par un vecteur viral.
Vecteurs viraux animaux et végétaux
Les virus qui infectent les cellules végétales et animales ont également été manipulés pour introduire des gènes étrangers dans les cellules végétales et animales. La capacité naturelle des virus à s'adsorber sur les cellules, à introduire leur ADN et à se répliquer en a fait des véhicules idéaux pour transférer l'ADN étranger dans les cellules eucaryotes en culture. Un vecteur basé sur le virus simien 40 (SV40) a été utilisé dans la première expérience de clonage impliquant des cellules de mammifères. Un certain nombre de vecteurs basés sur d'autres types de virus comme les adénovirus et le virus du papillome ont été utilisés pour cloner des gènes chez les mammifères. À l'heure actuelle, les vecteurs rétroviraux sont populaires pour le clonage de gènes dans les cellules de mammifères. Dans le cas des plantes, comme le virus de la mosaïque du chou-fleur , le virus de la mosaïque du tabac et les virus Gemini ont été utilisés avec un succès limité.
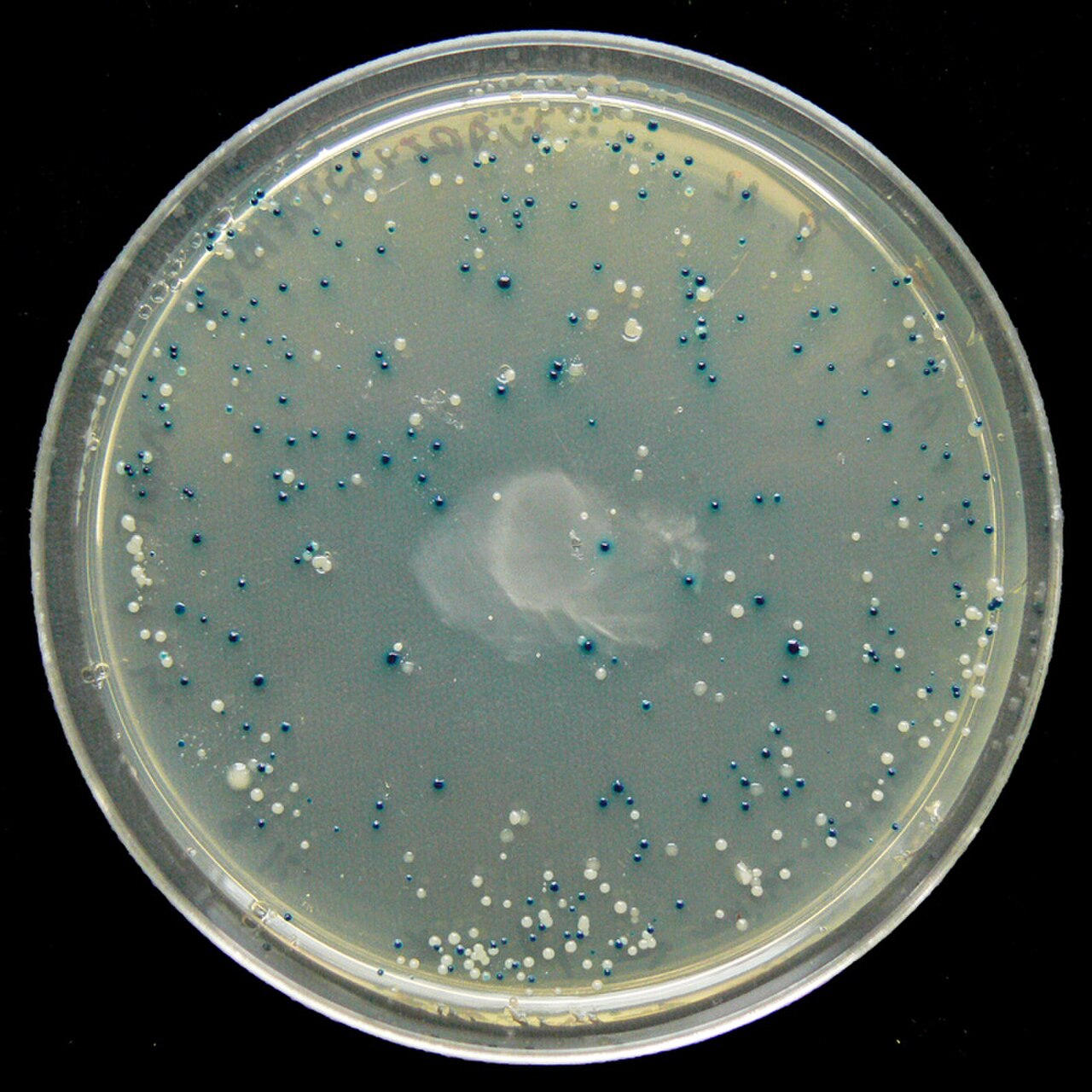
Dépistage : exemple de l'écran bleu/blanc
De nombreux vecteurs à usage général tels que pUC19 incluent généralement un système de détection de la présence d'un fragment d'ADN cloné, basé sur la perte d'un phénotype facilement détectable. Le gène le plus largement utilisé est le gène codant pour la β-galactosidase d' E. coli , dont l'activité peut être facilement détectée par la capacité de l'enzyme qu'il code à hydrolyser le substrat soluble et incolore X-gal (5-bromo-4-chloro-3-indolyl-bêta-d-galactoside) en un produit bleu insoluble (5,5'-dibromo-4,4'-dichloro indigo). Le clonage d'un fragment d'ADN dans la séquence lacZα basée sur le vecteur de la β-galactosidase empêche la production d'une enzyme active. Si X-gal est inclus dans les plaques d'agar sélectives, les colonies transformantes sont généralement bleues dans le cas d'un vecteur sans ADN inséré et blanches dans le cas d'un vecteur contenant un fragment d'ADN cloné.