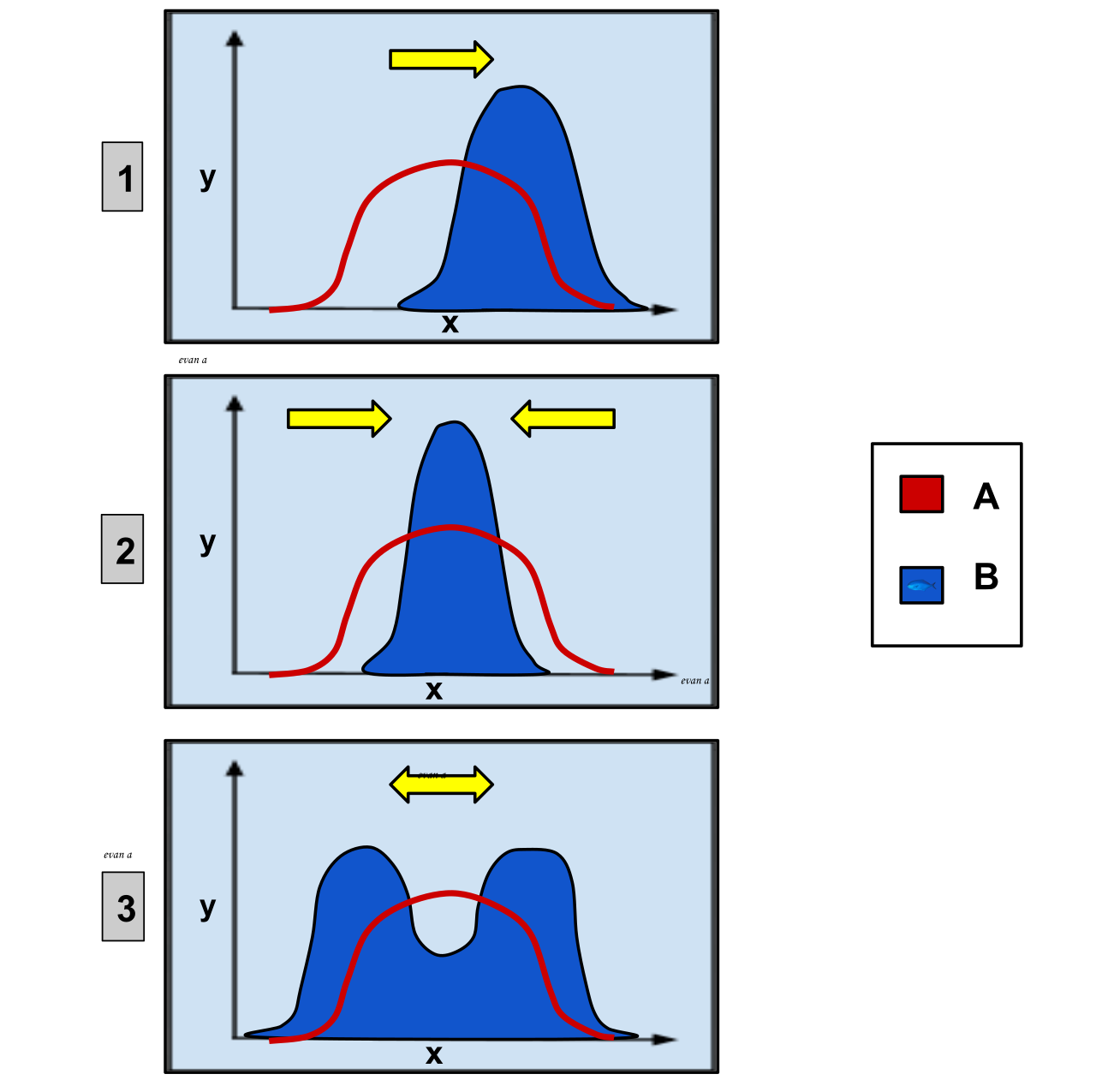
En biologie évolutive , la sélection disruptive , également appelée sélection diversifiante , décrit des changements dans la génétique des populations dans lesquels les valeurs extrêmes d'un trait sont favorisées par rapport aux valeurs intermédiaires. Dans ce cas, la variance du trait augmente et la population est divisée en deux groupes distincts. Dans ce cas, davantage d'individus acquièrent une valeur de caractère périphérique aux deux extrémités de la courbe de distribution.
Aperçu
La sélection naturelle est connue pour être l'un des processus biologiques les plus importants à l'origine de l'évolution . Il existe de nombreuses variations de traits, et certains entraînent un succès reproductif plus ou moins élevé de l'individu. L'effet de la sélection est de promouvoir certains allèles , traits et individus qui ont une plus grande chance de survivre et de se reproduire dans leur environnement spécifique. Étant donné que l'environnement a une capacité de charge , la nature agit sur ce mode de sélection sur les individus pour ne laisser que la progéniture la plus apte survivre et se reproduire à son plein potentiel. Plus le trait est avantageux, plus il deviendra courant dans la population. La sélection disruptive est un type spécifique de sélection naturelle qui sélectionne activement contre l'intermédiaire dans une population, favorisant les deux extrêmes du spectre.
On suppose que la sélection disruptive conduit souvent à une spéciation sympatrique par le biais d'un mode d'évolution par gradualisme phylétique . La sélection disruptive peut être causée ou influencée par de multiples facteurs et avoir également de multiples conséquences, en plus de la spéciation. Les individus au sein d'un même environnement peuvent développer une préférence pour les extrêmes d'un trait, par rapport au trait intermédiaire. La sélection peut agir sur des morphologies corporelles divergentes pour accéder à la nourriture, comme le bec et la structure dentaire. On constate que cela est souvent plus fréquent dans les environnements où il n'y a pas une large gamme clinique de ressources, ce qui entraîne un désavantage pour les hétérozygotes ou une sélection favorisant les homozygotes.
La répartition des niches permet de sélectionner des modèles différentiels d'utilisation des ressources, ce qui peut favoriser la spéciation. En revanche, la conservation des niches attire les individus vers des traits écologiques ancestraux dans un bras de fer évolutif. De plus, la nature a tendance à adopter une perspective de « saut dans le train en marche » lorsqu'elle découvre quelque chose d'utile. Cela peut conduire à l'inverse, avec une sélection disruptive qui finit par sélectionner contre la moyenne ; lorsque tout le monde commence à tirer parti de cette ressource, elle s'épuise et les extrêmes sont favorisés. De plus, le gradualisme est une vision plus réaliste de la spéciation par rapport à l'équilibre ponctué.
La sélection disruptive peut initialement intensifier rapidement la divergence, car elle ne manipule que les allèles déjà existants. Souvent, elle ne crée pas de nouveaux allèles par mutation, ce qui prend beaucoup de temps. En général, l'isolement reproductif complet ne se produit pas avant plusieurs générations, mais des différences comportementales ou morphologiques empêchent généralement les espèces de se reproduire. De plus, les hybrides ont généralement une fitness réduite, ce qui favorise l'isolement reproductif.
Exemple
Supposons qu'il existe une population de lapins. La couleur des lapins est régie par deux traits incomplètement dominants : la fourrure noire, représentée par « B », et la fourrure blanche, représentée par « b ». Un lapin de cette population avec un génotype « BB » aurait un phénotype de fourrure noire, un génotype « Bb » aurait une fourrure grise (une combinaison de noir et de blanc), et un génotype « bb » aurait une fourrure blanche.
Si cette population de lapins vivait dans un environnement comportant des zones de roches noires ainsi que des zones de roches blanches, les lapins à fourrure noire pourraient se cacher des prédateurs parmi les roches noires, et les lapins à fourrure blanche de même parmi les roches blanches. Les lapins à fourrure grise, en revanche, se distingueraient dans toutes les zones de l'habitat et subiraient ainsi une plus grande prédation.
En conséquence de ce type de pression sélective, notre population hypothétique de lapins serait sélectionnée de manière disruptive pour des valeurs extrêmes du trait de couleur de la fourrure : blanc ou noir, mais pas gris. Il s'agit d'un exemple de sous-dominance (désavantage hétérozygote) conduisant à une sélection disruptive.
Spéciation sympatrique
On pense que la sélection disruptive est l'une des principales forces qui conduisent à la spéciation sympatrique dans les populations naturelles. Les voies qui mènent de la sélection disruptive à la spéciation sympatrique sont rarement sujettes à des déviations ; une telle spéciation est un effet domino qui dépend de la cohérence de chaque variable distincte. Ces voies sont le résultat d'une sélection disruptive dans la compétition intraspécifique ; elle peut provoquer un isolement reproductif et finalement aboutir à la spéciation sympatrique.
Il est important de garder à l’esprit que la sélection disruptive ne doit pas toujours être basée sur la compétition intraspécifique. Il est également important de savoir que ce type de sélection naturelle est similaire aux autres. Lorsqu’elle n’est pas le facteur majeur, la compétition intraspécifique peut être écartée lors de l’évaluation des aspects opérationnels du processus d’adaptation. Par exemple, ce qui peut motiver la sélection disruptive plutôt que la compétition intraspécifique pourrait être des polymorphismes qui conduisent à l’isolement reproductif, et donc à la spéciation.
Lorsque la sélection disruptive est basée sur la compétition intraspécifique, la sélection qui en résulte favorise à son tour la diversification des niches écologiques et les polymorphismes. Si plusieurs morphes ( formes phénotypiques ) occupent des niches différentes, on peut s'attendre à ce que cette séparation favorise une concurrence réduite pour les ressources. La sélection disruptive est plus souvent observée dans les populations à haute densité que dans les populations à faible densité, car la concurrence intraspécifique tend à être plus intense au sein des populations à plus forte densité. En effet, les populations à plus forte densité impliquent souvent une plus grande concurrence pour les ressources. La concurrence qui en résulte pousse les polymorphismes à exploiter différentes niches ou des changements de niches afin d'éviter la concurrence. Si une morphe n'a pas besoin des ressources utilisées par une autre morphe, il est probable qu'aucune des deux ne subira de pression pour rivaliser ou interagir, favorisant ainsi la persistance et éventuellement l'intensification de la distinction des deux morphes au sein de la population. Cette théorie n'a pas nécessairement beaucoup de preuves à l'appui dans les populations naturelles, mais elle a été observée à de nombreuses reprises dans des situations expérimentales utilisant des populations existantes. Ces expériences confirment que, dans les situations appropriées (comme celles décrites ci-dessus), cette théorie pourrait s'avérer vraie dans la nature.
Lorsque la compétition intraspécifique n'est pas à l'œuvre, la sélection disruptive peut toujours conduire à la spéciation sympatrique et elle le fait en maintenant les polymorphismes. Une fois les polymorphismes maintenus dans la population, si l'accouplement assortatif a lieu, alors c'est une façon dont la sélection disruptive peut conduire à la spéciation sympatrique. Si différentes morphes ont des préférences d'accouplement différentes, alors l'accouplement assortatif peut se produire, surtout si le trait polymorphe est un « trait magique », c'est-à-dire un trait qui est soumis à la sélection écologique et qui a à son tour un effet secondaire sur le comportement reproductif. Dans une situation où le trait polymorphe n'est pas un trait magique, il doit y avoir une sorte de pénalité de fitness pour les individus qui ne s'accouplent pas de manière assortative et un mécanisme qui provoque l'accouplement assortatif doit évoluer dans la population. Par exemple, si une espèce de papillons développe deux types de motifs d'ailes, essentiels à des fins de mimétisme dans leur habitat préféré, alors l'accouplement entre deux papillons de motifs d'ailes différents conduit à un hétérozygote défavorable . Par conséquent, les papillons auront tendance à s'accoupler avec d'autres papillons ayant le même motif d'ailes, ce qui favorise une meilleure condition physique, éliminant finalement complètement l'hétérozygote. Cet hétérozygote défavorable génère une pression pour un mécanisme qui provoque l'accouplement assortatif qui conduira ensuite à un isolement reproductif en raison de la production de barrières post-accouplement. Il est en fait assez courant d'observer une spéciation sympatrique lorsque la sélection disruptive soutient deux morphes, en particulier lorsque le trait phénotypique affecte la condition physique plutôt que le choix du partenaire .
Dans les deux situations, l’une où la compétition intraspécifique est à l’œuvre et l’autre où elle ne l’est pas, si tous ces facteurs sont réunis, ils conduiront à un isolement reproductif, qui peut conduire à une spéciation sympatrique.
Autres résultats
- polymorphisme
- dimorphisme sexuel
- plasticité phénotypique
Importance
La sélection disruptive revêt une importance particulière dans l'histoire de l'étude de l'évolution, car elle est impliquée dans l'un des « cas cardinaux » de l'évolution, à savoir les populations de pinsons observées par Darwin aux Galápagos . Il a observé que les espèces de pinsons étaient suffisamment similaires pour être apparemment issues d'une seule espèce. Cependant, elles présentaient une variation disruptive de la taille du bec. Cette variation semblait être liée de manière adaptative à la taille des graines disponibles sur les îles respectives (gros becs pour les grosses graines, petit bec pour les petites graines). Les becs moyens avaient du mal à récupérer les petites graines et n'étaient pas non plus assez résistants pour les plus grosses graines, ce qui les rendait inadaptés.
S’il est vrai que la sélection disruptive peut conduire à la spéciation, ce processus n’est pas aussi rapide ou simple que d’autres types de spéciation ou de changement évolutif. Cela introduit le sujet du gradualisme, qui est une accumulation lente mais continue de changements sur de longues périodes de temps. Cela est dû en grande partie au fait que les résultats de la sélection disruptive sont moins stables que ceux de la sélection directionnelle (la sélection directionnelle ne favorise que les individus situés à une extrémité du spectre).
Prenons par exemple le cas mathématiquement simple mais biologiquement improbable des lapins : supposons qu'une sélection directionnelle ait lieu. Le champ ne contient que des pierres sombres, donc plus le lapin est sombre, plus il peut se cacher efficacement des prédateurs. Il finira par y avoir beaucoup de lapins noirs dans la population (donc beaucoup d'allèles « B ») et une quantité moindre de lapins gris (qui contribuent à 50 % des chromosomes avec l'allèle « B » et à 50 % des chromosomes avec l'allèle « b » à la population). Il y aura peu de lapins blancs (peu de contributeurs de chromosomes avec l'allèle « b » à la population). Cela pourrait finalement conduire à une situation dans laquelle les chromosomes avec l'allèle « b » s'éteignent, faisant du noir la seule couleur possible pour tous les lapins suivants. La raison en est qu'il n'y a rien qui « augmente » le niveau de chromosomes « b » dans la population. Ils ne peuvent que diminuer, et finalement s'éteindre.
Considérons maintenant le cas de la sélection disruptive. Le résultat est un nombre égal de lapins noirs et blancs, et donc un nombre égal de chromosomes avec l'allèle « B » ou « b », qui flottent toujours dans cette population. Chaque fois qu'un lapin blanc s'accouple avec un lapin noir, seuls des lapins gris en résultent. Ainsi, pour que les résultats soient cohérents, il faut qu'une force pousse les lapins blancs à choisir d'autres lapins blancs et les lapins noirs à choisir d'autres lapins noirs. Dans le cas des pinsons, cette « force » était l'isolement géographique/de niche. Cela nous amène à penser que la sélection disruptive ne peut pas se produire et qu'elle est normalement due à l'isolement géographique des espèces, à la sélection directionnelle ou à la sélection stabilisatrice.