Un complexe de jonction d'exons ( EJC ) est un complexe protéique qui se forme sur un brin d'ARN pré-messager à la jonction de deux exons qui ont été joints ensemble lors de l'épissage de l'ARN . L'EJC a des influences majeures sur la traduction , la surveillance , la localisation de l' ARNm épissé et la méthylation m6A . Il est d'abord déposé sur l'ARNm lors de l'épissage, puis transporté dans le cytoplasme . Il y joue un rôle majeur dans la régulation post-transcriptionnelle de l'ARNm. On pense que les complexes de jonction d'exons fournissent une mémoire spécifique de position de l'événement d'épissage. L'EJC se compose d'un noyau hétérotétramère stable, qui sert de plate-forme de liaison pour d'autres facteurs nécessaires à la voie de l'ARNm. Le cœur de l'EJC contient la protéine facteur d'initiation eucaryote 4A-III ( eIF4A-III ; une hélicase à ARN à boîte DEAD ) liée à un analogue de l'adénosine triphosphate ( ATP ), ainsi qu'aux protéines supplémentaires Magoh et Y14 . La liaison de ces protéines aux domaines mouchetés du noyau a été mesurée récemment et elle peut être régulée par les voies de signalisation PI3K/AKT/mTOR . Pour que la liaison du complexe à l'ARNm se produise, le facteur eIF4AIII est inhibé, ce qui arrête l'hydrolyse de l'ATP. Cela reconnaît l'EJC comme un complexe dépendant de l'ATP. L'EJC interagit également avec un grand nombre de protéines supplémentaires ; notamment les protéines SR. Ces interactions sont considérées comme importantes pour la compaction de l'ARNm. Le rôle de l'EJC dans l'exportation de l'ARNm est controversé.
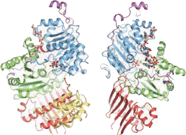
Composants protéiques
Français L'EJC est composé de plusieurs composants protéiques clés : RNPS1 , Y14, SRm160 , Aly/REF et Magoh, entre autres. RNPS1 peut fonctionner comme un coactivateur de l'épissage, mais avec Y14, il participe également au processus de dégradation induite par le non-sens chez les eucaryotes. SRm160 est un autre coactivateur qui a été proposé pour améliorer le traitement de l'extrémité 3' de l'ARNm. Le composant protéique Magoh est censé faciliter la localisation sous-cytoplasmique des ARNm tandis qu'Aly est engagé dans l'exportation de l'ARNm nucléaire. On pense qu'Aly est recruté dans le complexe de jonction d'exons par la protéine UAP56 . UAP56 est reconnu comme une hélicase à ARN mais agit comme un facteur d'épissage nécessaire à l'assemblage précoce du spliceosome. Un autre facteur impliqué dans la voie EJC est DEK . Ce composant est connu pour participer à une variété de fonctions allant de l'épissage à la régulation transcriptionnelle et à la structure de la chromatine .
Structure
La cristallisation du complexe de jonction exonique a révélé l'organisation structurelle de ses composants protéiques. Le noyau du complexe est allongé avec une dimension globale de 99 Å par 67 Å par 54 Å. Il est organisé autour du facteur eIF4AIII. Le facteur lui-même se compose de deux types de conformations différentes autour de l'ARNm : fermé et ouvert. Dans un état fermé, les deux domaines d'eIF4AIII forment des sites de liaison composites pour le 5'-adénylyl-β,γ-imidodiphosphate (ADPNP) et l'ARNm. Dans la conformation ouverte, les deux domaines sont tournés de 160 degrés par rapport à l'état fermé. Les composants protéiques Magoh et Y14 se lient ensemble pour former un hétérodimère situé au pôle 5' de l'EJC. Magoh se lie à un domaine eIF4AIII par des interactions entre les résidus de ses deux hélices C-terminales et une extrémité d'une grande feuille β . Les résidus conservés dans le lieur entre les deux domaines eIF4AIII forment des ponts salins ou des liaisons hydrogène avec des résidus spécifiques de Magoh. D'autres liaisons se produisent entre la deuxième boucle de la feuille β de Magoh et les deux domaines eIF4AIII et leur lieur. Il n'y a qu'une seule liaison partielle formée entre Y14 et eIF4AIII. Il s'agit d'un pont salin entre les résidus conservés Y14 Arg108 et eIF4AIII Asp401 . Si des mutations devaient se produire sur ces deux résidus, l'association de Magoh-Y14 avec EJC serait inexistante.
Mécanisme
Au cours de la deuxième étape de l'épissage dans les cellules eucaryotes, l'EJC est déposé à environ 20-24 nucléotides de l'extrémité 5' en amont de la jonction d'épissage (où deux exons sont joints), lorsque le lariat s'est formé et que les exons sont ligaturés ensemble. La liaison de l'EJC à l'ARNm se produit de manière indépendante de la séquence, pour former la ribonucléoprotéine messagère mature (mRNP). L'EJC reste lié de manière stable à cette mRNP lorsqu'elle est exportée hors du noyau et dans le cytoplasme. Les composants protéiques sont liés ou libérés par l'EJC lors de son transport. Pour que la translocation des ARNm à travers le complexe des pores nucléaires se produise, un hétérodimère composé de NXF1 /TAP et NXT1 / p15 doit se lier aux transcriptions. NXF1/TAP est un récepteur majeur pour l'exportation des ARNm vers le cytoplasme. Cela est dû au fait qu'il interagit à la fois avec les protéines adaptatrices de liaison à l'ARN et avec les composants du complexe des pores nucléaires .
La reconnaissance d'un codon de terminaison prématurée se produit lors de la traduction dans le cytoplasme. L'image ci-dessous suggère que cet événement est nucléaire, contrairement à l'opinion générale dans ce domaine. Les lecteurs doivent être conscients que la traduction dans le noyau est un sujet très controversé qui n'est pas bien étayé par des données.
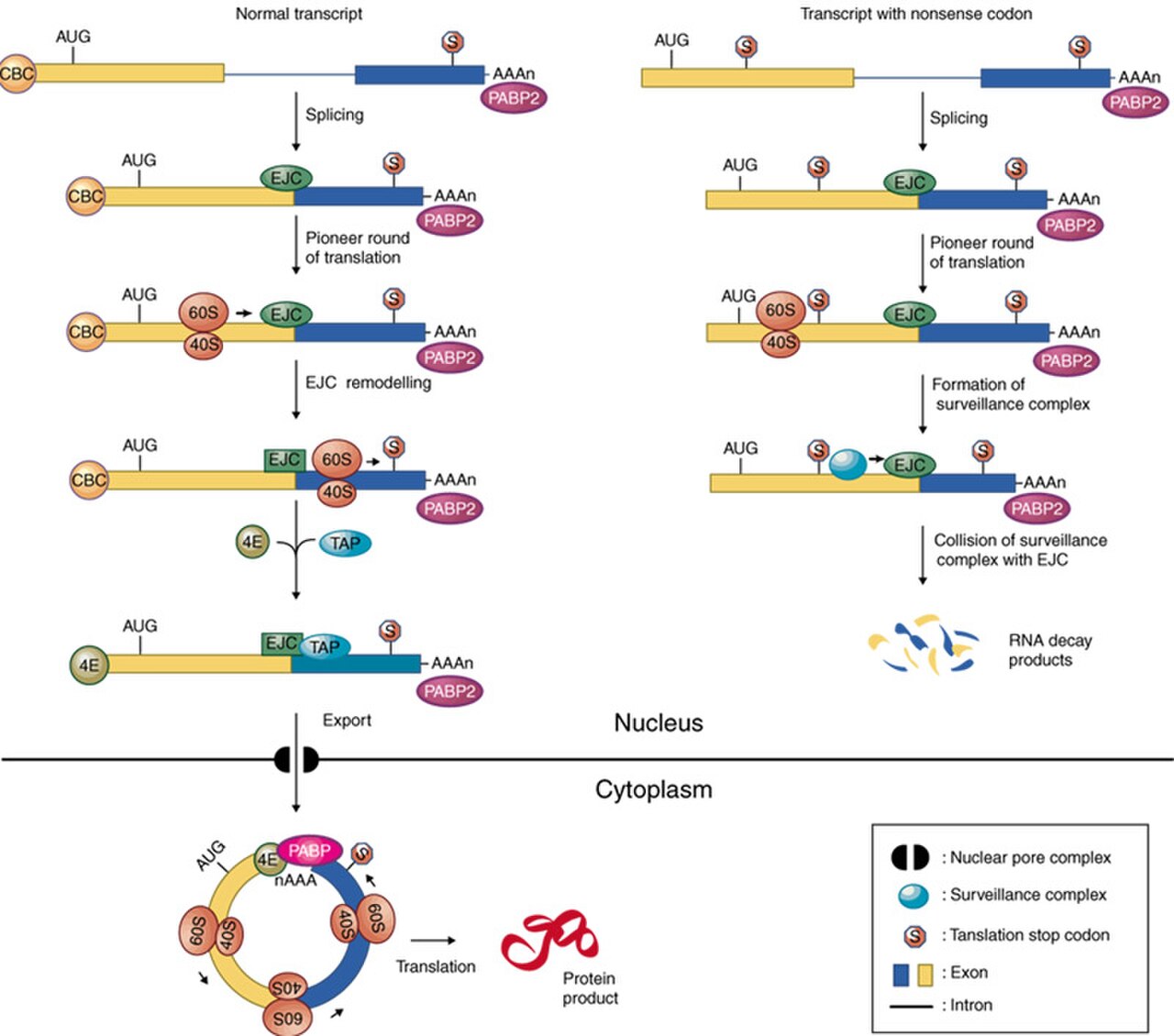
Dans la décadence médiatisée par le non-sens
Les complexes de jonction exonique jouent un rôle majeur dans la surveillance de l'ARNm . Plus précisément, ils se trouvent dans la voie de dégradation médiée par le non-sens (NMD), dans laquelle les transcrits d'ARNm avec des codons d'arrêt prématurés sont dégradés. Dans la traduction normale de l'ARNm, le ribosome se lie au transcrit et commence l'élongation de la chaîne d'acides aminés . Il continue jusqu'à ce qu'il atteigne l'emplacement du complexe de jonction exonique, qu'il déplace ensuite. Ensuite, la traduction est terminée lorsque le ribosome atteint un codon de terminaison . Dans la NMD, le transcrit d'ARNm contient un codon de terminaison prématuré (PTC) en raison d'une mutation non-sens . Si ce codon se produit avant le site EJC, l'EJC restera lié, déclenchant la dégradation de l'ARNm. L'EJC et sa position servent de type de régulateur, déterminant si le transcrit est défectueux ou non.
Les EJC sont également connus pour participer au NMD d'une autre manière ; le recrutement des facteurs de surveillance UPF1 , UPF2 et UPF3 . Ces protéines sont les composants les plus importants du mécanisme NMD. Les protéines EJC MAGOH, Y14 et eIF4AIII assurent la liaison à UPF3, qui agit comme un pont entre UPF2 et UPF1 en formant un complexe trimérique. Au sein de ce complexe, UPF2 et UPF3 agissent en coopération pour promouvoir l'ATPase et l'hélicase à ARN de UPF1. Le noyau EJC ancre de manière stable le complexe UPF à l'ARNm et aide à la régulation de la protéine essentielle UPF1. Les ribosomes qui sont bloqués sur un PTC recrutent UPF1 par le biais d'interactions avec le facteur de libération eRF1 et eRF3 . Avec la protéine SMG1 , eRF1, eRF3 et UPF1 forment le complexe SURF. Ce complexe forme un pont entre le ribosome et l'EJC en aval qui est associé à UPF3 et UPF2. Cette interaction déclenche la phosphorylation de UPF1 par SMG1, provoquant la dissociation de eRF1 et eRF3. Le complexe produit se compose d'EJC, UPF3, UPF2, UPF1 phosphorylé et SMG1 et déclenche à son tour la dégradation de l'ARNm.