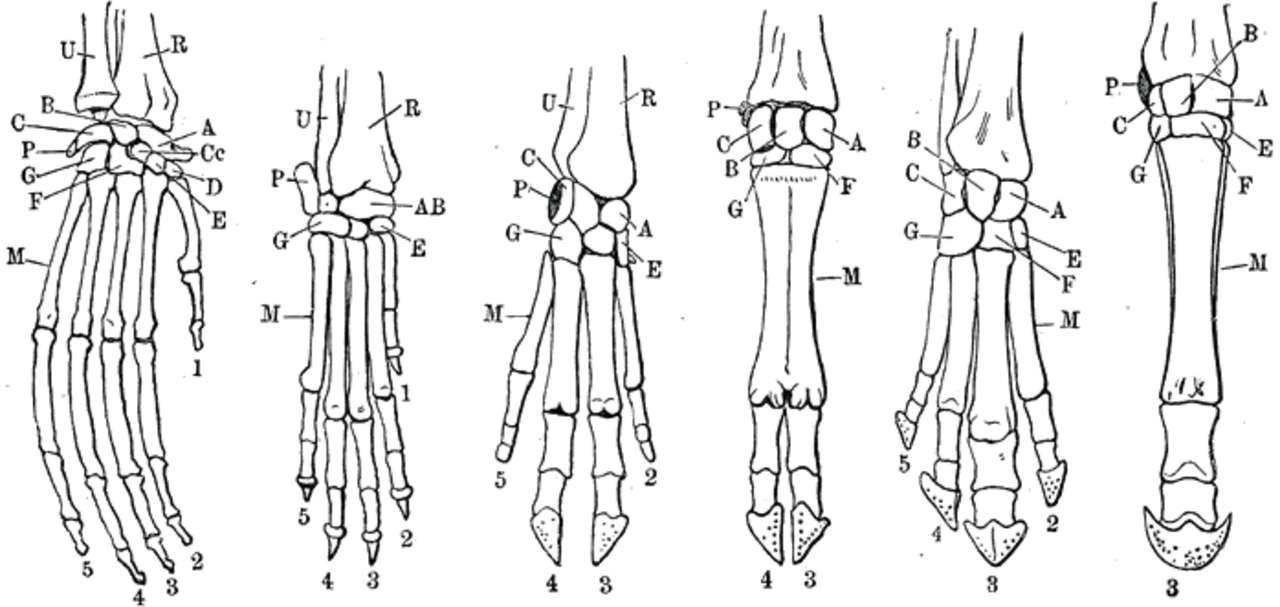
Un membre antérieur ou membre antérieur est l'un des appendices articulés appariés ( membres ) fixés à l' extrémité crânienne ( antérieure ) du torse d' un vertébré tétrapode terrestre . En référence aux quadrupèdes , le terme patte antérieure ou membre antérieur est souvent utilisé à la place. Chez les animaux bipèdes ayant une posture verticale (par exemple les humains et certains primates ), le terme membre supérieur est souvent utilisé.
Un membre antérieur ne doit pas être confondu avec un avant-bras , qui est une partie distale du membre supérieur humain entre le coude et le poignet .
Tous les membres antérieurs des vertébrés sont homologues , ce qui signifie qu'ils ont tous évolué à partir des mêmes structures. Par exemple, la nageoire d'une tortue ou d'un dauphin , le bras d'un humain, la patte avant d'un cheval et les ailes des chauves-souris et des oiseaux sont finalement homologues, malgré les grandes différences qui les existent entre eux.
Les utilisations spécifiques des membres antérieurs peuvent être analogues si elles ont évolué à partir de différentes sous-structures du membre antérieur, telles que les nageoires des tortues et des dauphins, et les ailes des oiseaux et des chauves-souris.
Evolution des membres antérieurs
L'évolution des membres antérieurs peut être caractérisée par de nombreuses tendances. Le nombre de doigts , leurs caractéristiques, ainsi que la forme et l'alignement du radius , du cubitus et de l'humérus ont eu des implications évolutives majeures.
On constate fréquemment que les changements dans la taille du corps, la posture des pieds, l’habitat et le substrat s’influencent mutuellement (et se connectent à des facteurs potentiels plus larges, tels que le changement climatique).
Forme
Plusieurs facteurs peuvent influencer l'évolution de la forme des os longs des membres antérieurs, comme la masse corporelle, le mode de vie, le comportement prédateur ou la taille relative des proies. En général, les espèces plus lourdes ont des radius, des cubitus et des humérus plus robustes.
Les carnivores mustéloïdes qui ont un mode de vie arboricole ont tendance à avoir des os longs et fins au niveau des membres antérieurs, ce qui permet une meilleure mobilité et une meilleure flexibilité. Les espèces mustéloïdes semi-fouisseuses et aquatiques ont tendance à avoir des os longs courts et robustes au niveau des membres antérieurs pour faire face à la tension due au creusement et à la nage.
Dans l'ordre des carnivores , les félins, qui tendent généralement des embuscades et luttent avec leurs proies, ont des membres plus courts et plus robustes. Leurs membres antérieurs sont utilisés à la fois pour les sprints courts et pour lutter, ce qui signifie qu'ils doivent être flexibles et résistants. En revanche, les canidés, qui poursuivent souvent leurs proies sur de plus grandes distances, ont des membres plus longs et plus graciles. La course est à peu près la seule utilité de leurs membres antérieurs, ils n'ont donc pas besoin d'être adaptés à autre chose et peuvent être moins flexibles.
Les prédateurs chassant des proies pesant la moitié de leur poids corporel ou plus ont développé des rayons, des cubitus et des humérus plus courts et plus robustes pour réduire le risque de fracture ou de rupture de l'os pendant la chasse. Les prédateurs chassant des proies pesant moins de la moitié de leur poids corporel avaient tendance à avoir des os longs des membres antérieurs plus longs et plus fins pour améliorer l'efficacité énergétique.
Polydactylie
On pensait initialement que les tétrapodes avaient d'abord développé cinq doigts comme caractéristique ancestrale, qui ont ensuite été réduits ou spécialisés dans un certain nombre d'utilisations. Certains animaux ont conservé des membres antérieurs « primitifs », comme les reptiles pentadactyles (à cinq doigts) et les primates. Cela s'est avéré en grande partie vrai, mais les premiers ancêtres tétrapodes ou « fishapodes » ont peut-être eu plus de cinq doigts. Cette hypothèse a été notamment contestée par Stephen Jay Gould dans son essai de 1991 « Eight (Or Fewer) Little Piggies »
La polydactylie chez les premiers tétrapodes doit être comprise comme la présence de plus de cinq doigts au niveau des doigts ou des pieds, une condition qui était la situation naturelle des tout premiers tétrapodes. Les premiers groupes comme Acanthostega avaient huit doigts, tandis que le plus dérivé Ichthyostega en avait sept, et le Tulerpeton encore plus dérivé en avait six.
Les tétrapodes ont évolué à partir d'animaux dotés de nageoires, comme celles que l'on trouve chez les poissons à nageoires lobées . À partir de cette condition, un nouveau modèle de formation des membres a évolué, où l'axe de développement du membre a tourné pour faire germer des axes secondaires le long de la marge inférieure, donnant naissance à un nombre variable de supports squelettiques très robustes pour un pied en forme de pagaie.
Spécialisation numérique
Les doigts peuvent être spécialisés pour différentes formes de locomotion. Un exemple classique est le développement d'un seul orteil chez le cheval (monodactylie). D'autres sabots, comme ceux des ongulés à doigts pairs et impairs , et même le pied en forme de sabot des hadrosaures éteints , peuvent être considérés comme des spécialisations similaires.
Pour supporter leur poids immense, les sauropodes , dont les plus dérivés sont les titanosaures , ont développé un manus tubulaire (pied avant) et ont progressivement perdu leurs doigts, se tenant debout sur leurs métacarpes. Le membre antérieur du stégosaure présente des preuves d'une configuration métacarpienne semblable à celle des sauropodes . Il s'agissait d'une stratégie évolutive différente de celle des mammifères mégafauniques tels que les éléphants modernes.
Les Thérapsides ont commencé à développer des membres antérieurs diversifiés et spécialisés il y a 270 millions d'années, au cours du Permien.
Pouce opposable
Les humains modernes sont uniques en ce qui concerne la musculature de l’avant-bras et de la main, bien que des pouces opposables ou des structures similaires soient apparus chez quelques animaux.
Chez les dinosaures, une autonomisation primitive de la première articulation carpométacarpienne (CMC) a pu se produire. Chez les primates, une véritable différenciation est apparue il y a peut-être 70 millions d'années, tandis que la forme de la CMC du pouce humain apparaît finalement il y a environ 5 millions d'années.
- Les primates se répartissent en quatre groupes :
- Pouces non opposables : tarsiers et ouistitis
- Pouces pseudo-opposables : tous les strepsirrhines et Cebidae
- Pouces opposables : singes de l'Ancien Monde et tous les grands singes
- Opposables avec des pouces relativement longs : gibbons (ou petits singes)
Les pandas ont développé des pouces pseudo-opposables par extension de l' os sésamoïde , qui n'est pas un vrai doigt.
Pronation et supination
La pronation de la main et de l'avant-bras chez les mammifères thériens est obtenue grâce à la tête arrondie du radius, qui lui permet de pivoter par rapport à l'ulna. La supination nécessite un glissement dorsal du radius distal et la pronation un glissement palmaire par rapport à l'ulna distal.
La pronation a évolué à plusieurs reprises, chez les mammifères , les caméléons et les varanides . Cependant, la condition la plus basique est de ne pas pouvoir se pronation. Les dinosaures n'étaient pas capables de faire plus que la semi-pronation du poignet, bien que les origines bipèdes de tous les clades de dinosaures quadrupèdes aient pu permettre une plus grande disparité dans la posture des membres antérieurs que ce que l'on pense souvent. Les monotrèmes ont des avant-bras qui ne sont pas aussi adroits que ceux des thériens. Les monotrèmes ont une posture tentaculaire et de multiples éléments dans leurs ceintures pectorales, qui sont des traits ancestraux pour les mammifères.
Chez les oiseaux, les muscles de l'avant-bras effectuent la supination, la pronation, la flexion et l'extension de l'aile distale.
Ailes
Les membres antérieurs de tous les tétrapodes sont homologues, évoluant à partir des mêmes structures initiales chez les poissons à nageoires lobées . Cependant, un autre processus distinct peut être identifié, l'évolution convergente , par laquelle les ailes des oiseaux , des chauves-souris et des ptérosaures éteints ont évolué vers le même but de manière radicalement différente. Ces structures ont une forme ou une fonction similaire mais n'étaient pas présentes chez le dernier ancêtre commun de ces groupes.
Les ailes des chauves-souris sont en grande partie constituées d'une fine membrane de peau soutenue par les cinq doigts, tandis que les ailes des oiseaux sont en grande partie constituées de plumes soutenues par des doigts beaucoup plus petits, le doigt 2 soutenant l' alula et le doigt 4 les plumes primaires de l'aile ; il n'y a que des homologies lointaines entre les oiseaux et les chauves-souris, avec des homologies beaucoup plus proches entre n'importe quelle paire d'espèces d'oiseaux, ou n'importe quelle paire d'espèces de chauves-souris.
Palmes
Les mammifères marins ont évolué à plusieurs reprises. Au cours de leur évolution, ils développent des corps hydrodynamiques profilés. Le membre antérieur se développe ainsi en nageoire . Les membres antérieurs des cétacés, des pinnipèdes et des siréniens présentent un exemple classique d'évolution convergente. Il existe une convergence généralisée au niveau des gènes. Des substitutions distinctes dans des gènes communs ont créé diverses adaptations aquatiques, dont la plupart constituent une évolution parallèle car les substitutions en question ne sont pas propres à ces animaux.
En comparant les cétacés aux pinnipèdes et aux siréniens, 133 substitutions parallèles d'acides aminés se produisent. En comparant et en contrastant les cétacés-pinnipèdes, les cétacés-siréniens et les pinnipèdes-siréniens, 2 351, 7 684 et 2 579 substitutions se produisent respectivement.