L'évolution parallèle est le développement similaire d'un trait chez des espèces distinctes qui ne sont pas étroitement liées, mais qui partagent un trait original similaire en réponse à une pression évolutive similaire.
Evolution parallèle vs. évolution convergente
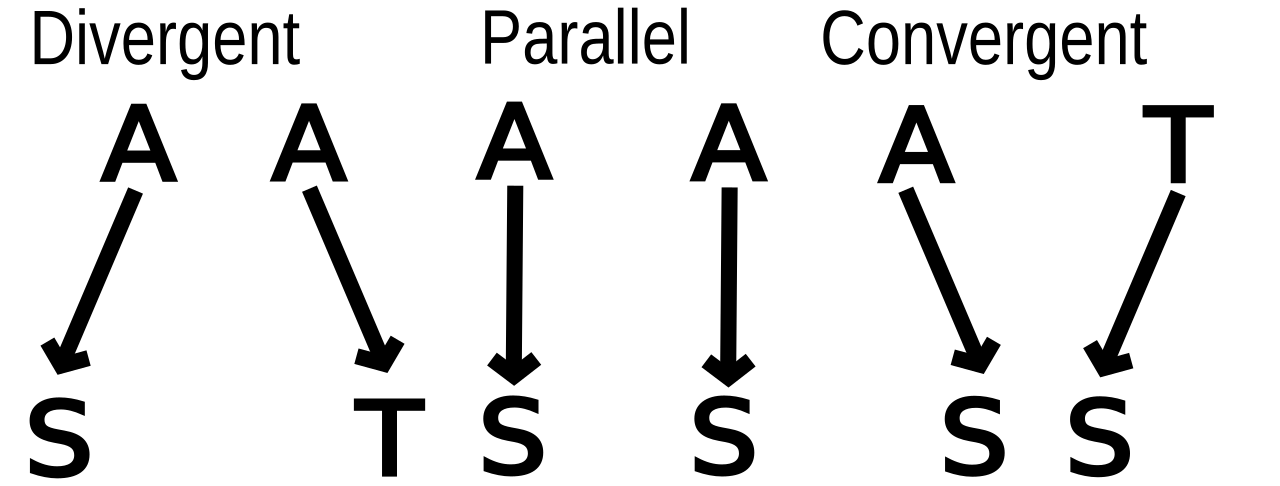
Étant donné un trait qui se produit dans chacune des deux lignées descendant d'un ancêtre spécifié, il est théoriquement possible de définir de manière stricte les tendances évolutives parallèles et convergentes et de les distinguer clairement les unes des autres. Cependant, les critères permettant de définir l'évolution convergente par opposition à l'évolution parallèle ne sont pas clairs dans la pratique, de sorte que le diagnostic arbitraire est courant. Lorsque deux espèces partagent un trait, l'évolution est définie comme parallèle si l'on sait que les ancêtres ont partagé cette similitude ; dans le cas contraire, elle est définie comme convergente. Cependant, les conditions énoncées sont une question de degré ; tous les organismes partagent des ancêtres communs. Les scientifiques diffèrent sur l'utilité de la distinction.
Évolution parallèle entre les marsupiaux et les placentaires
Plusieurs exemples d'évolution parallèle sont fournis par les deux principales branches des mammifères , les placentaires et les marsupiaux , qui ont suivi des voies évolutives indépendantes après la fragmentation des masses continentales telles que le Gondwana il y a environ 100 millions d'années. En Amérique du Sud , les marsupiaux et les placentaires partageaient l'écosystème (avant le Grand Échange Américain ) ; en Australie , les marsupiaux prévalaient ; et dans l' Ancien Monde et en Amérique du Nord, les placentaires l'emportaient. Cependant, dans toutes ces localités, les mammifères étaient de petite taille et n'occupaient qu'une place limitée dans l'écosystème jusqu'à l' extinction massive des dinosaures il y a 65 millions d'années. À cette époque, les mammifères des trois masses continentales ont commencé à adopter une plus grande variété de formes et de rôles. Si certaines formes étaient propres à chaque environnement, des animaux étonnamment similaires ont souvent émergé sur deux ou trois des continents séparés. Parmi ces espèces, on peut citer les chats à dents de sabre placentaires ( Machairodontinae ) et le marsupial à dents de sabre d'Amérique du Sud ( Thylacosmilus ) ; le loup de Tasmanie et le loup européen ; de même que les taupes marsupiales et placentaires , les écureuils volants et (sans doute) les souris .
Coévolution parallèle des traits entre les colibris et les souimangas contribuant aux guildes écologiques
Les colibris et les souimangas, deux lignées d'oiseaux nectarivores du Nouveau et de l'Ancien Monde, ont développé en parallèle une série de traits comportementaux et anatomiques spécialisés . Ces traits (forme du bec, enzymes digestives et vol ) permettent aux oiseaux de s'adapter de manière optimale à la niche écologique de pollinisation et d'alimentation des fleurs qu'ils occupent, qui est façonnée par les séries de traits parallèles des oiseaux. Ainsi, un syndrome comportemental coévolutif parallèle au sein des oiseaux crée une guilde émergente d'oiseaux hautement spécialisés et de plantes hautement adaptées, chacun exploitant l'implication de l'autre dans la pollinisation des fleurs dans l'Ancien et le Nouveau Monde.
La forme du bec des nectarivores, long et en forme d'aiguille, leur permet d'atteindre le pistil/l'étamine d'une fleur et d'atteindre le nectar qu'il contient. Les nectarivores peuvent également utiliser leur bec spécialisé pour se livrer au vol de nectar , une pratique observée à la fois chez les colibris et les souimangas dans laquelle l'oiseau obtient du nectar en faisant un trou dans la base du tube de la corolle de la fleur au lieu d'insérer son bec dans le tube comme c'est la norme, « volant » ainsi le nectar de la fleur puisqu'elle n'est pas pollinisée en retour.
Les nectarivores et les fleurs ornithophiles existent souvent dans des relations de guilde mutualistes facilitées par la forme du bec de l'oiseau, la source de nourriture et la capacité digestive agissant de concert avec la forme du tube de la fleur et l'adaptation à la pollinisation par les oiseaux en vol stationnaire ou perchés. Les oiseaux mangent du nectar à l'aide de leur bec long et fin et, ce faisant, collectent du pollen sur leur bec ; ce pollen est ensuite transféré à la fleur suivante dont ils se nourrissent. Ce mutualisme a coévolué en parallèle entre les oiseaux de l'Ancien et du Nouveau Monde et leurs fleurs respectives. De plus, l'activité enzymatique digestive des nectarivores correspondant à la composition du nectar de leurs fleurs respectives semble avoir coévolué en parallèle entre les plantes et les pollinisateurs à travers les continents, car les lignées nectarivores ont développé indépendamment la capacité de digérer le nectar spécifique à leurs fleurs, ce qui a donné lieu à des guildes distinctes.
La capacité des nectarivores à digérer le saccharose est bien supérieure à celle des autres taxons aviaires . Cette différence est due à une concentration analogue élevée de sucrase-isomaltase , une enzyme qui hydrolyse le saccharose. L'activité de la sucrase par unité de surface intestinale semble être plus élevée chez les nectarivores que chez les autres oiseaux, ce qui signifie que ces oiseaux nectarivores peuvent digérer plus de saccharose plus rapidement que les autres taxons. De plus, l'hypothèse de modulation adaptative ne s'applique pas aux nectarivores et aux enzymes de digestion du sucre, ce qui signifie que deux lignées de nectarivores ne devraient pas nécessairement avoir toutes deux des concentrations élevées de sucrase-isomaltase même si elles mangent toutes deux du nectar. Ainsi, l'acquisition parallèle d'une capacité digestive analogue du saccharose est une conclusion raisonnable car il n'y a aucune raison apparente pour que les deux lignées partagent cette concentration élevée enzymatique.