Représentation schématique de l'assemblage des histones du noyau en nucléosome En biologie , les histones sont des protéines basiques riches en résidus de lysine et d'arginine ,...
Worldlex WikiContenu en francaisLecture gratuite
Représentation schématique de l'assemblage des histones du noyau en nucléosome
En biologie , les histones sont des protéines basiques riches en résidus de lysine et d'arginine , présentes dans les noyaux des cellules eucaryotes et chez la plupart des archées . Elles agissent comme des bobines autour desquelles l'ADN s'enroule pour former des unités structurales appelées nucléosomes . Les nucléosomes s'enroulent ensuite en fibres de 30 nanomètres qui constituent la chromatine , un réseau compact de molécules d'ADN . Les histones empêchent l'ADN de s'enchevêtrer et le protègent des dommages . De plus, elles jouent un rôle important dans la régulation des gènes et la réplication de l'ADN . Sans histones, l'ADN déroulé des chromosomes serait très long. Par exemple, chaque cellule humaine possède environ 1,8 mètre d'ADN s'il était complètement étiré ; cependant, enroulé autour des histones, cette longueur est réduite à environ 9 micromètres (0,009 mm) de fibres de chromatine de 30 nm de diamètre.
Il existe cinq familles d'histones, désignées H1/H5 (histones de liaison), H2, H3 et H4 (histones du nucléosome). Le nucléosome est constitué de deux dimères H2A-H2B et d'un tétramère H3-H4 . L'enroulement étroit de l'ADN autour des histones résulte en grande partie de l'attraction électrostatique entre les histones, chargées positivement, et le squelette phosphate de l'ADN, chargé négativement.
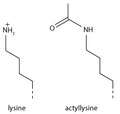
Les histones peuvent être modifiées chimiquement par l'action d'enzymes afin de réguler la transcription des gènes. Les modifications les plus fréquentes sont la méthylation des résidus d'arginine ou de lysine, ou l' acétylation de la lysine. La méthylation peut influencer l'interaction d'autres protéines, telles que les facteurs de transcription, avec les nucléosomes. L'acétylation de la lysine élimine sa charge positive, affaiblissant ainsi l'attraction électrostatique entre l'histone et l'ADN. Il en résulte un déroulement partiel de l'ADN, le rendant plus accessible à l'expression des gènes.
Hétérooctamère d'histones (H3, H4, H2A, H2B) + fragment d'ADN, grenouille
Il existe cinq grandes familles de protéines histones : H1/H5 , H2A , H2B , H3 et H4 . Les histones H2A, H2B, H3 et H4 sont connues sous le nom d'histones de cœur ou nucléosomales, tandis que les histones H1/H5 sont connues sous le nom d'histones de liaison .
Les histones du noyau existent toutes sous forme de dimères , qui présentent la particularité de posséder le domaine de repliement des histones : trois hélices alpha reliées par deux boucles. C’est cette structure hélicoïdale qui permet l’interaction entre les différents dimères, notamment selon un mécanisme tête-queue (également appelé motif de poignée de main). Les quatre dimères distincts ainsi formés s’assemblent ensuite pour former un noyau nucléosomique octamérique d’environ 63 angströms de diamètre (une particule de type solénoïde (ADN) ). Environ 146 paires de bases (pb) d’ADN s’enroulent autour de ce noyau 1,65 fois en un tour superhélicoïdal gauche, donnant une particule d’environ 100 angströms de diamètre. L’histone de liaison H1 se lie au nucléosome aux sites d’entrée et de sortie de l’ADN, stabilisant ainsi ce dernier et permettant la formation de structures d’ordre supérieur. La plus simple de ces structures est la conformation en fibre de 10 nm ou en collier de perles. Ce processus implique l'enroulement de l'ADN autour des nucléosomes , chaque paire de nucléosomes étant séparée par environ 50 paires de bases d' ADN (également appelées ADN de liaison ). Parmi les structures d'ordre supérieur, on trouve la fibre de 30 nm (formant un zigzag irrégulier) et la fibre de 100 nm, présentes dans les cellules normales. Lors de la mitose et de la méiose, les chromosomes condensés s'assemblent grâce aux interactions entre les nucléosomes et d'autres protéines régulatrices.
Les histones sont subdivisées en histones canoniques dépendantes de la réplication, dont les gènes sont exprimés durant la phase S du cycle cellulaire , et en variants d'histones indépendants de la réplication , exprimés tout au long du cycle cellulaire. Chez les mammifères, les gènes codant pour les histones canoniques sont généralement regroupés le long des chromosomes en quatre loci différents et hautement conservés , sont dépourvus d'introns et possèdent une structure en tige-boucle à leur extrémité 3' au lieu d'une queue polyA . Les gènes codant pour les variants d'histones ne sont généralement pas regroupés, possèdent des introns et leurs ARNm sont régulés par des queues polyA. Les organismes multicellulaires complexes présentent généralement un plus grand nombre de variants d'histones, assurant diverses fonctions. Sur le plan fonctionnel, les variants d'histones contribuent au contrôle transcriptionnel, à la mémoire épigénétique et à la réparation de l'ADN, remplissant des fonctions spécialisées au-delà du compactage des nucléosomes, qui joue un rôle distinct dans la dynamique de la chromatine. Par exemple, H2A.Z est enrichi au niveau des éléments régulateurs et des promoteurs des gènes activement transcrits, où il module la stabilité des nucléosomes et la liaison des facteurs de transcription. En revanche, H3.3, un variant de remplacement de l'histone H3 , est associé à la transcription active et se dépose préférentiellement au niveau des éléments amplificateurs et des corps de gènes transcrits. Un autre variant essentiel, CENPA , remplace H3 dans les nucléosomes centromériques, fournissant une base structurale indispensable à la ségrégation chromosomique.
Les variants jouent également un rôle essentiel dans la réparation de l'ADN . Des variants tels que H2A.X sont phosphorylés au niveau des lésions de l'ADN, marquant ainsi des régions cibles pour le recrutement des protéines de réparation. Cette modification, communément appelée γH2A.X, constitue un signal clé dans la réponse cellulaire aux cassures double brin , facilitant ainsi des processus de réparation de l'ADN efficaces. Des anomalies dans la régulation des variants d'histones ont été associées à l'instabilité génomique , une caractéristique de nombreux cancers et maladies liées à l'âge.
Des données récentes s'accumulent concernant les rôles des différentes variantes d'histones, mettant en évidence les liens fonctionnels entre ces variantes et la régulation fine du développement des organismes. Les protéines variantes d'histones de différents organismes, leur classification et leurs caractéristiques spécifiques sont répertoriées dans la base de données « HistoneDB 2.0 - Variants » . Plusieurs pseudogènes ont également été découverts et identifiés dans des séquences très proches de leurs gènes orthologues fonctionnels respectifs.
Voici une liste des protéines histones humaines, des gènes et des pseudogènes :
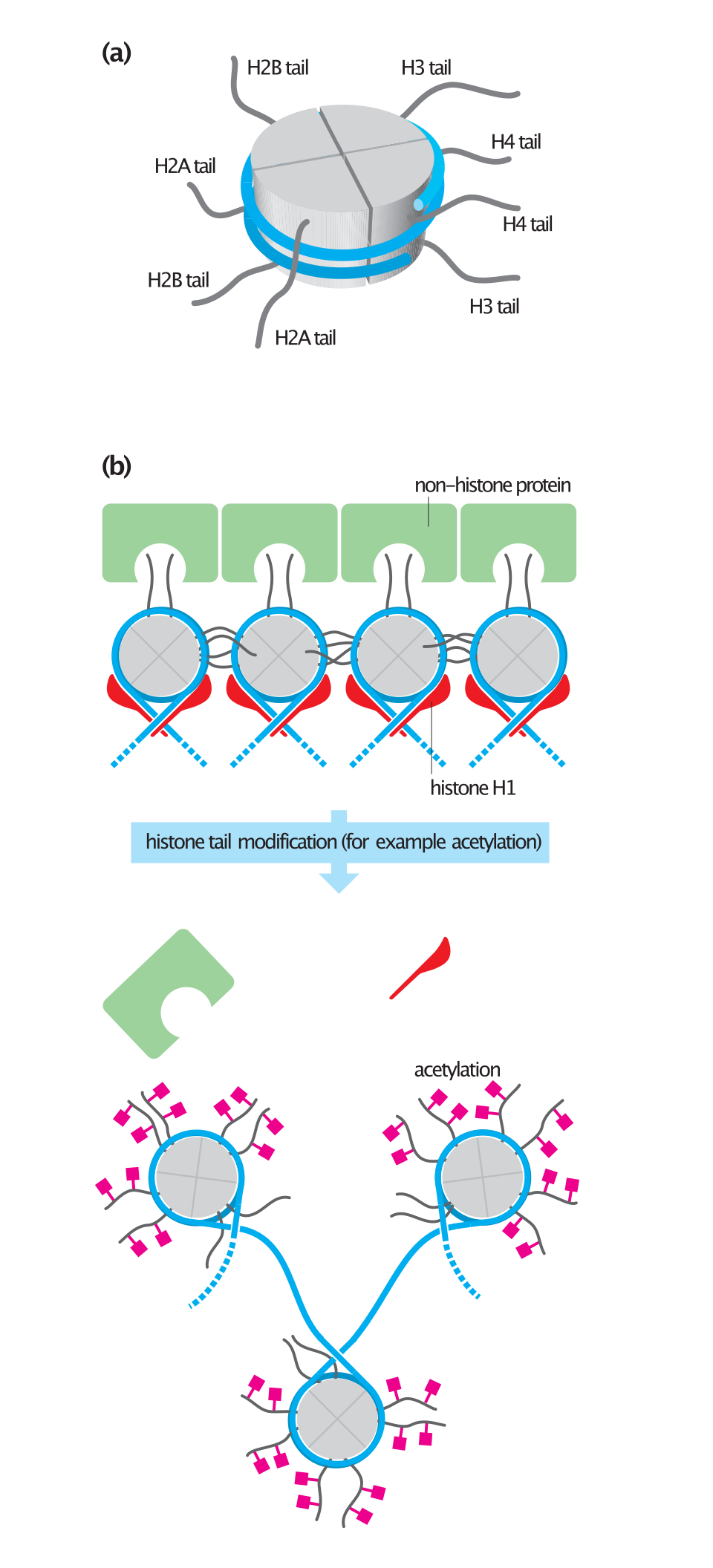
Le cœur du nucléosome est formé de deux dimères H2A-H2B et d'un tétramère H3-H4, formant deux moitiés quasi symétriques par leur structure tertiaire ( symétrie C2 ; une macromolécule est l'image miroir de l'autre). Les dimères H2A-H2B et le tétramère H3-H4 présentent également une symétrie pseudodyadique. Les quatre histones « cœur » (H2A, H2B, H3 et H4) sont relativement similaires et hautement conservées au cours de l'évolution . Elles possèdent toutes un motif « hélice-tour -hélice -tour-hélice » (motif de liaison à l'ADN qui reconnaît une séquence d'ADN spécifique). Elles partagent également la caractéristique de longues « queues » à une extrémité de leur structure en acides aminés , site de modifications post-traductionnelles (voir ci-dessous).
La distance entre les bobines autour desquelles les cellules eucaryotes enroulent leur ADN a été déterminée comme étant comprise entre 59 et 70 Å.
Au total, les histones établissent cinq types d'interactions avec l'ADN :
Ponts salins et liaisons hydrogène entre les chaînes latérales des acides aminés basiques (en particulier la lysine et l'arginine ) et les atomes d'oxygène des phosphates de l'ADN
Les dipôles hélice-hélice forment des hélices alpha dans H2B, H3 et H4, ce qui entraîne une accumulation de charge positive nette au point d'interaction avec les groupes phosphate chargés négativement sur l'ADN.
Liaisons hydrogène entre le squelette de l'ADN et le groupe amide de la chaîne principale des protéines histones
Interactions non polaires entre les histones et les désoxyriboses sur l'ADN
Insertion non spécifique des queues N-terminales des histones H3 et H2B dans deux sillons mineurs de la molécule d'ADN
La nature hautement basique des histones, outre le fait qu'elle facilite les interactions ADN-histones, contribue à leur solubilité dans l'eau.
En général, les gènes actifs présentent une moindre association avec les histones, tandis que les gènes inactifs y sont fortement associés durant l'interphase . Il apparaît également que la structure des histones a été conservée au cours de l'évolution , car toute mutation délétère serait gravement inadaptée. Toutes les histones possèdent une extrémité N-terminale fortement chargée positivement et riche en résidus de lysine et d'arginine .
Au-delà des eucaryotes
Les histones archéennes présentent une organisation beaucoup plus diversifiée, bien qu'elles ne contiennent que l'équivalent des repliements des histones centrales. Elles suivent des voies d'évolution des histones centrales que les eucaryotes n'ont pas nécessairement empruntées. La plupart des histones archéennes s'assemblent en une structure dimérique de type H3-H4, composée d'un seul type d'unité. Ces structures dimériques peuvent s'empiler pour former une grande superhélice (« hypernucléosome ») sur laquelle l'ADN s'enroule à la manière des nucléosomes, comme démontré chez Thermococcus et Methanothermus . 33 % des génomes archéens ne possèdent pas de génome d'histones identifiable et 28 % ne possèdent qu'un seul gène de type H3/H4, sans la spécialisation en quatre types principaux observée chez les eucaryotes. Les autres, en revanche, présentent un certain degré de spécialisation, notamment des histones inhabituelles du point de vue eucaryote.
Des analyses bioinformatiques ont également identifié des histones archéennes acides. Les simulations de dynamique moléculaire (DM) suggèrent qu'elles conservent leur capacité à enrouler l'ADN autour de structures semblables à des nucléosomes grâce au caractère basique de la « crête » en contact avec l'ADN. Plus inhabituel est le repliement double des histones , constitué de deux repliements d'histones reliés par un segment de liaison au sein d'une même chaîne protéique, plus fréquent chez les archées halophiles. Les simulations de DM suggèrent que si ces histones ne peuvent enrouler l'ADN autour de structures semblables à des nucléosomes à faible concentration saline, elles en sont capables à forte concentration. Parmi les autres types inhabituels, on trouve l'histone « miniature » acide (deuxième hélice tronquée) et le repliement quadruple des histones.
Les Nucleocytoviricota (grands virus à ADN nucléocytoplasmiques, NCLDV) comprennent également des exemples d'histones. Ils possèdent des représentants des quatre groupes d'histones centrales eucaryotes (H2A, H2B, H3, H4). Certains virus contiennent les quatre, parfois fusionnées en une seule protéine. Ils s'auto-assemblent en nucléosomes de type eucaryote qui s'empilent en oligomères de type archéen. Compte tenu de cette caractéristique intermédiaire et du point de divergence dans les arbres phylogénétiques, ils pourraient avoir divergé des eucaryotes au cours de l'eucaryogenèse .
Des protéines similaires aux histones du noyau sont également présentes chez une minorité de bactéries, probablement en raison d'un transfert horizontal de gènes. Deux exemples sont caractérisés : Bdellovibrio bacteriovorus utilise un mode « par la tranche » qui ne forme pas de nucléosomes (mais cela est controversé) et noyaux des cellules eucaryotes et chez la plupart des archées, mais pas chez les bactéries . On pensait auparavant que les dinoflagellés, algues unicellulaires, étaient les seuls eucaryotes totalement dépourvus d'histones , mais des études ultérieures ont montré que leur ADN code pour des gènes d'histones . Contrairement aux histones nucléocellulaires, des homologues des protéines histones de liaison riches en lysine (H1) sont présents chez les bactéries, sous le nom de nucléoprotéines HC1/HC2
Il a été suggéré que les protéines histones centrales sont apparentées, d'un point de vue évolutif, à la partie hélicoïdale du domaine ATPase AAA+ étendu (domaine C) et au domaine de reconnaissance du substrat N-terminal des protéines Clp/Hsp100. Malgré leurs différences topologiques, ces trois structures partagent un motif homologue hélice-brin-hélice (HSH) . Il est également proposé qu'elles pourraient avoir évolué à partir de protéines ribosomiques ( RPS6 / RPS15 ), ces deux dernières étant des protéines courtes et basiques
Les histones archéennes pourraient bien ressembler aux précurseurs évolutifs des histones eucaryotes. Les protéines histones figurent parmi les protéines les plus conservées chez les eucaryotes, soulignant leur rôle important dans la biologie du noyau. En revanche, les spermatozoïdes matures utilisent principalement des protamines pour compacter leur ADN génomique, probablement parce que cela leur permet d'atteindre un taux de compaction encore plus élevé.
Il existe des variantes au sein de certaines classes majeures d'histones. Elles partagent une homologie de séquence en acides aminés et une similarité structurale centrale avec une classe spécifique d'histones majeures, mais possèdent également des caractéristiques qui leur sont propres et distinctes de celles des histones majeures. Ces histones mineures assurent généralement des fonctions spécifiques du métabolisme de la chromatine. Par exemple, l'histone CENPA , apparentée à l'histone H3, est associée exclusivement à la région centromérique du chromosome. La variante H2A.Z de l'histone H2A est associée aux promoteurs des gènes activement transcrits et participe également à la prévention de la propagation de l' hétérochromatine silencieuse . De plus, H2A.Z joue un rôle dans la stabilité du génome au sein de la chromatine . Une autre variante de H2A, H2A.X, est phosphorylée en S139 dans les régions entourant les cassures double brin et marque la région en cours de réparation de l'ADN . L'histone H3.3 est associée au corps des gènes activement transcrits
FonctionUnités de base de la structure de la chromatine
Compactage des brins d'ADN
Les histones agissent comme des bobines autour desquelles s'enroule l'ADN. Ceci permet la compaction nécessaire pour loger les grands génomes des eucaryotes dans les noyaux cellulaires : la molécule compactée est 40 000 fois plus courte qu'une molécule non compactée.
Transcription
L’effet des histones sur la transcription a été établi dès la fin des années 1980. Plus précisément, lorsque les histones sont mutées ou épuisées, une activation substantielle de la transcription se produit et est indépendante des autres facteurs de transcription (ou suggérant que ces derniers déplacent les histones).
Régulation de la chromatine
Les queues d'histones et leur fonction dans la formation de la chromatine
ModificationReprésentation schématique des modifications des histones. D'après Rodriguez-Paredes et Esteller, Nature, 2011.
Un vaste catalogue de modifications d'histones a été établi, mais la plupart d'entre elles restent encore mal comprises sur le plan fonctionnel. On pense généralement que ces modifications pourraient constituer la base d'un code des histones , où des combinaisons de modifications auraient des significations spécifiques. Cependant, la plupart des données fonctionnelles concernent des modifications d'histones individuelles importantes, susceptibles d'être étudiées biochimiquement en détail.
Chimie
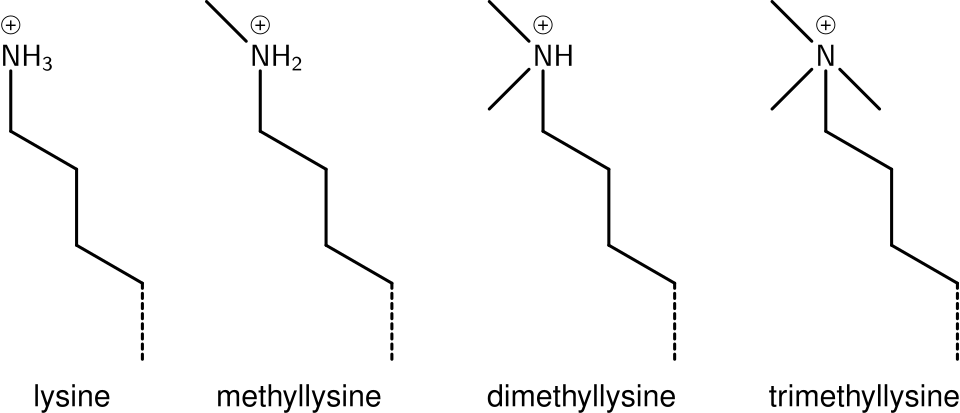
Méthylation de la lysine
L'ajout d'un, deux ou plusieurs groupements méthyle à la lysine a peu d'effet sur la chimie de l'histone ; la méthylation préserve la charge de la lysine et n'ajoute qu'un nombre minimal d'atomes, de sorte que les interactions stériques restent globalement inchangées. Cependant, les protéines contenant des domaines Tudor, chromo ou PHD, entre autres, peuvent reconnaître la méthylation de la lysine avec une sensibilité remarquable et différencier les lysines mono-, di- et triméthylées, au point que, pour certaines lysines (par exemple : H4K20), ces trois formes de méthylation semblent avoir des significations distinctes. De ce fait, la méthylation de la lysine constitue un marqueur très informatif et prédomine parmi les fonctions connues de modification des histones.
sérotonine à la glutamine en position 5 de l'histone H3 se produit dans les cellules sérotoninergiques, notamment les neurones. Ce processus participe à la différenciation de ces cellules. Cette modification post-traductionnelle est concomitante à la modification H3K4me3. La sérotoninylation potentialise la liaison du facteur de transcription général TFIID à la boîte TATA .
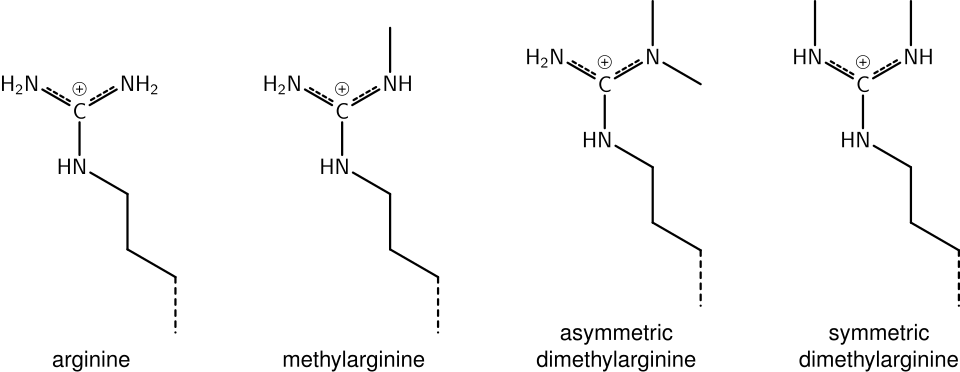
méthylation de l'arginine
Ce qui a été dit précédemment concernant la chimie de la méthylation de la lysine s'applique également à la méthylation de l'arginine, et certains domaines protéiques, comme les domaines Tudor, peuvent être spécifiques de la méthylarginine plutôt que de la méthyllysine. L'arginine est connue pour être mono- ou diméthylée, et la méthylation peut être symétrique ou asymétrique, avec potentiellement différentes significations.
citrullination de l'arginine
Les enzymes appelées peptidylarginine désiminases (PAD) hydrolysent le groupe imine des arginines et y fixent un groupe céto, ce qui diminue la charge positive du résidu d'acide aminé. Ce processus participe à l'activation de l'expression génique en rendant les histones modifiées moins fortement liées à l'ADN et, par conséquent, la chromatine plus accessible. Les PAD peuvent également produire l'effet inverse en supprimant ou en inhibant la monométhylation des résidus d'arginine sur les histones, et ainsi antagoniser l'effet positif de la méthylation de l'arginine sur l'activité transcriptionnelle.
Acétylation de la lysine
L'ajout d'un groupe acétyle a un effet chimique majeur sur la lysine, car il neutralise sa charge positive. Ceci réduit l'attraction électrostatique entre l'histone et le squelette de l'ADN, chargé négativement, ce qui relâche la structure de la chromatine ; les histones fortement acétylées forment une chromatine plus accessible et sont généralement associées à une transcription active. L'acétylation de la lysine semble moins précise que la méthylation, car les histones acétyltransférases agissent souvent sur plusieurs lysines ; ceci reflète probablement la nécessité de modifier plusieurs lysines pour avoir un effet significatif sur la structure de la chromatine. Cette modification inclut H3K27ac .
L'ajout d'un groupe phosphate chargé négativement peut induire des modifications importantes de la structure des protéines, ce qui explique le rôle bien établi de la phosphorylation dans la régulation de leur fonction. Les implications structurales de la phosphorylation des histones restent à élucider, mais ses fonctions en tant que modification post-traductionnelle sont clairement définies, et des domaines de liaison tels que BRCT ont été caractérisés.
Effets sur la transcription
Les modifications des histones interviennent dans le contrôle de la transcription.
Gènes activement transcrits
Deux modifications d'histones sont particulièrement associées à la transcription active :
Triméthylation de la lysine 4 de l'histone H3 (H3K4me3)
Cette triméthylation se produit au niveau du promoteur des gènes actifs et est catalysée par le complexe COMPASS . Malgré la conservation de ce complexe et de cette modification d'histone de la levure aux mammifères, le rôle précis de cette modification reste encore mal défini. Elle constitue néanmoins un excellent marqueur des promoteurs actifs, et le niveau de cette modification d'histone au niveau du promoteur d'un gène est globalement corrélé à l'activité transcriptionnelle de ce gène. La formation de cette marque est liée à la transcription de manière complexe : au début de la transcription d'un gène, l'ARN polymérase II passe d'une phase d' initiation à une phase d'élongation , caractérisée par une modification de l'état de phosphorylation du domaine C-terminal (CTD) de l'ARN polymérase II . La même enzyme qui phosphoryle le CTD phosphoryle également le complexe Rad6 , lequel ajoute à son tour une marque d'ubiquitine à H2B K123 (K120 chez les mammifères) . H2BK123Ub est présent dans toutes les régions transcrites, mais cette marque est nécessaire à la triméthylation de H3K4 par COMPASS au niveau des promoteurs
Triméthylation de la lysine 36 de l'histone H3 ( H3K36me3 )
Cette triméthylation se produit au sein des gènes actifs et est catalysée par la méthyltransférase Set2 . Cette protéine s'associe à l'ARN polymérase II en cours d'élongation , et la présence de H3K36Me3 est caractéristique des gènes activement transcrits . H3K36Me3 est reconnue par le complexe histone désacétylase Rpd3, qui élimine les modifications acétylées des histones environnantes, augmentant ainsi la compaction de la chromatine et réprimant la transcription aberrante . L'augmentation de la compaction de la chromatine empêche les facteurs de transcription d'accéder à l'ADN et réduit la probabilité d'initiation de nouvelles transcriptions au sein du gène. Ce processus contribue donc à garantir la continuité de la transcription.
gènes réprimés
Trois modifications d'histones sont particulièrement associées aux gènes réprimés :
Triméthylation de la lysine 27 de l'histone H3 (H3K27me3)
Cette modification d'histone est déposée par le complexe polycomb PRC2 . Elle constitue un marqueur clair de la répression génique et est probablement liée à d'autres protéines pour exercer une fonction répressive. Un autre complexe polycomb , PRC1, peut se lier à H3K27me3 et ajoute la modification d'histone H2AK119Ub, qui favorise la compaction de la chromatine . D'après ces données, il semblerait que PRC1 soit recruté par l'intermédiaire de PRC2. Cependant, des études récentes montrent que PRC1 est recruté aux mêmes sites en l'absence de PRC2
Di- et triméthylation de la lysine 9 de l'histone H3 (H3K9me2/3)
Triméthylation de la lysine 20 de l'histone H4 ( H4K20me3 )
Cette modification est étroitement associée à l'hétérochromatine, bien que son importance fonctionnelle reste incertaine. Cette marque est déposée par la méthyltransférase Suv4-20h, qui est au moins en partie recrutée par la protéine d'hétérochromatine 1.
promoteurs bivalents
L'analyse des modifications d'histones dans les cellules souches embryonnaires (et autres cellules souches) a révélé de nombreux promoteurs de gènes portant à la fois les marques H3K4Me3 et H3K27Me3 ; autrement dit, ces promoteurs présentent simultanément des marques d'activation et de répression. Cette combinaison particulière de modifications caractérise les gènes prêts à être transcrits ; ces modifications ne sont pas nécessaires dans les cellules souches, mais le deviennent rapidement après la différenciation en certaines lignées. Une fois la différenciation cellulaire amorcée, ces promoteurs bivalents adoptent un état actif ou répressif selon la lignée choisie.
Autres fonctions
réparation des dommages à l'ADN
Le marquage des sites de lésions de l'ADN est une fonction importante des modifications d'histones. Sans marqueur de réparation, l'ADN serait détruit par les dommages accumulés, notamment ceux provenant du rayonnement ultraviolet du soleil.
Phosphorylation de H2AX à la sérine 139 (γH2AX)
L'histone H2AX phosphorylée (également appelée gamma H2AX) est un marqueur des cassures réponse aux dommages à l'ADN . La phosphorylation de H2AX survient précocement après la détection d'une cassure double brin de l'ADN et forme un domaine s'étendant sur plusieurs kilobases de part et d'autre de la lésion La gamma H2AX sert de site de liaison à la protéine MDC1, qui recrute à son tour des protéines clés de la réparation de l'ADN (ce sujet complexe est bien décrit dans ). De ce fait, la gamma H2AX constitue un élément essentiel du mécanisme garantissant la stabilité du génome.
Acétylation de la lysine 56 de l'histone H3 (H3K56Ac)
L'acétylation de H3K56 ( H3K56Acx) est essentielle à la stabilité du génome. H3K56 est acétylée par le complexe p300/Rtt109, mais est rapidement désacétylée au niveau des lésions de l'ADN. L'acétylation de H3K56 est également nécessaire à la stabilisation des fourches de réplication bloquées, empêchant ainsi leur effondrement dangereux. Bien que les mammifères utilisent généralement beaucoup plus les modifications d'histones que les micro-organismes, un rôle majeur de H3K56Ac dans la réplication de l'ADN n'est observé que chez les champignons, ce qui en fait une cible pour le développement d'antibiotiques.
Triméthylation de la lysine 36 de l'histone H3 (H3K36me3)
La méthylation H3K36me3 permet le recrutement du complexe MSH2-MSH6 (hMutSα) de la voie de réparation des mésappariements de l'ADN . De façon cohérente, les régions du génome humain présentant des niveaux élevés de H3K36me3 accumulent moins de mutations somatiques grâce à l'activité de réparation des mésappariements .
Condensation chromosomique
Phosphorylation de H3 à la sérine 10 (phospho-H3S10)
La kinase mitotique Aurora B phosphoryle l'histone H3 sur la sérine 10, déclenchant une cascade de modifications qui induisent la condensation des chromosomes mitotiques. Les chromosomes condensés présentent donc une forte coloration pour cette marque, mais la phosphorylation de H3S10 est également présente à certains endroits des chromosomes en dehors de la mitose, par exemple dans l'hétérochromatine péricentromérique des cellules en phase G2. La phosphorylation de H3S10 a également été associée à des lésions de l'ADN causées par la formation de boucles R au niveau de régions fortement transcrites.
Phosphorylation de H2B à la sérine 10/14 (phospho-H2BS10/14)
La phosphorylation de l'histone H2B sur la sérine 10 (levure) ou la sérine 14 (mammifères) est également liée à la condensation de la chromatine, mais dans un but très différent : celui de médier la condensation des chromosomes lors de l'apoptose. Cette marque n'est pas simplement un élément secondaire tardif de l'apoptose, car les levures porteuses de mutations de ce résidu sont résistantes à la mort cellulaire apoptotique induite par le peroxyde d'hydrogène.
Dépendance
Les modifications épigénétiques des queues d'histones dans des régions spécifiques du cerveau jouent un rôle central dans les addictions. Une fois apparues, ces altérations épigénétiques semblent constituer des « cicatrices moléculaires » durables qui pourraient expliquer la persistance des addictions.
Les fumeurs de cigarettes (environ 15 % de la population américaine) sont généralement dépendants à la nicotine . Après sept jours de traitement à la nicotine chez la souris, l'acétylation des histones H3 et H4 était accrue au niveau du promoteur de FosB dans le noyau accumbens , entraînant une augmentation de 61 % de l'expression de FosB. Ceci augmenterait également l'expression du variant d'épissage ΔFosB . Dans le noyau accumbens , ΔFosB agit comme un « interrupteur moléculaire durable » et une « protéine de contrôle principale » dans le développement de la dépendance .
Environ 7 % de la population américaine est dépendante à l'alcool . Chez des rats exposés à l'alcool pendant une durée allant jusqu'à 5 jours, on a observé une augmentation de l'acétylation de la lysine 9 de l'histone 3 dans le promoteur de la pronociceptine au niveau du complexe amygdalien . Cette acétylation constitue une marque d'activation de la pronociceptine. Le système nociceptine/récepteur opioïde de la nociceptine est impliqué dans les effets de renforcement ou de conditionnement de l'alcool.
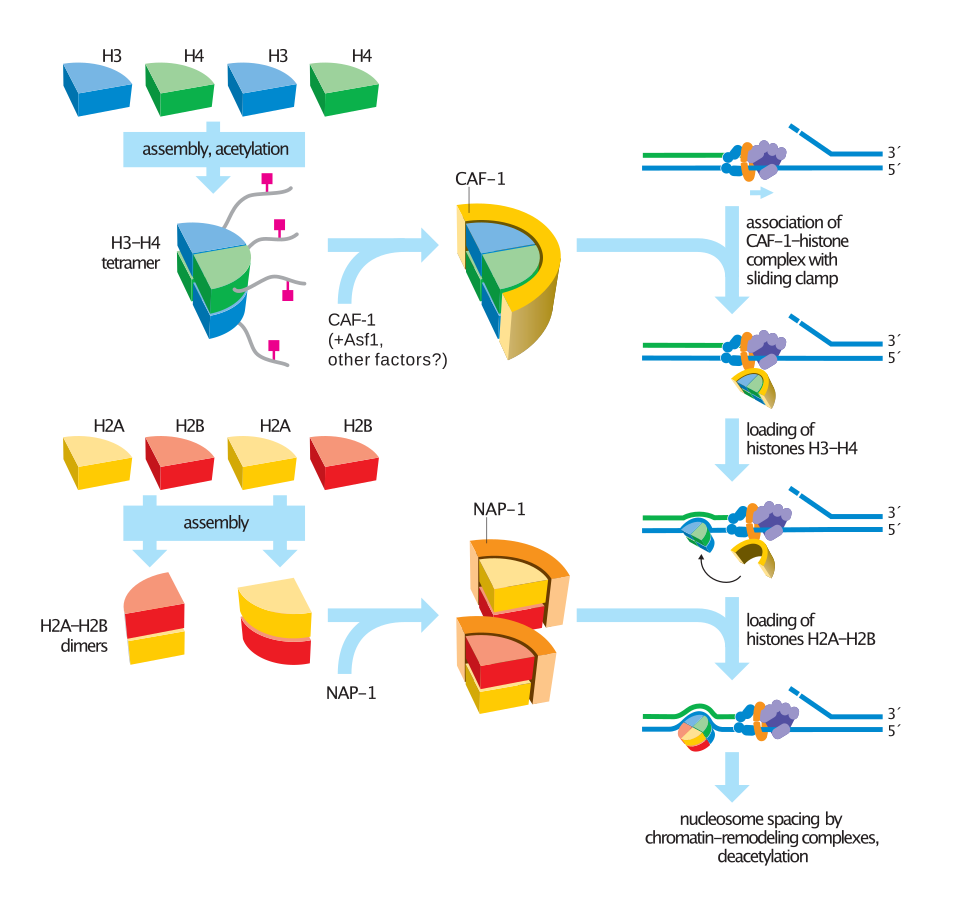
Les chaperonnes d'histones sont des protéines spécialisées qui participent à la manipulation, au transport et à l'assemblage corrects des histones, empêchant leur agrégation et assurant leur dépôt approprié sur l'ADN. Ces protéines jouent un rôle crucial dans la régulation de l'assemblage et du désassemblage des nucléosomes , influençant l'activité transcriptionnelle, la réplication et la réparation de l'ADN. Contrairement au remodelage enzymatique de la chromatine , les chaperonnes d'histones agissent en se liant aux histones de manière régulée, modulant ainsi la structure de la chromatine sans activité catalytique directe.
L'une des fonctions essentielles des chaperonnes d'histones est de maintenir un stock d'histones et de réguler leur apport afin de garantir une formation correcte de la chromatine. Lors de la réplication et de la transcription de l'ADN , des chaperonnes d'histones telles que ASF1 et FACT facilitent le réassemblage des nucléosomes, assurant ainsi la préservation des modifications d'histones qui définissent l'identité cellulaire. De plus, les chaperonnes d'histones contribuent au désassemblage des nucléosomes en réponse à un stress cellulaire ou à des lésions de l'ADN, permettant ainsi l'accès aux mécanismes de réparation.
Les chaperonnes d'histones participent également au dépôt sélectif de variants d'histones, fonctionnellement distincts des histones canoniques . Par exemple, HIRA est une chaperonne qui dépose spécifiquement le variant d'histone H3.3, un marqueur des régions de chromatine active. De même, CAF-1 est responsable de l'incorporation de H3.1 et H3.2 dans l'ADN nouvellement répliqué, ce qui souligne la spécialisation fonctionnelle au sein des réseaux de chaperonnes.
Compte tenu de leur rôle crucial, la dérégulation des chaperonnes d'histones est impliquée dans des maladies telles que le cancer. Une activité aberrante de ces chaperonnes peut entraîner un dépôt incorrect d'histones, une instabilité génomique et une altération de l'expression des gènes, contribuant ainsi à la tumorigénèse . Les recherches actuelles explorent les chaperonnes d'histones comme cibles thérapeutiques potentielles, notamment dans les cancers caractérisés par des perturbations de la chromatine.
réseaux d'accompagnateurs
L'action coordonnée de plusieurs chaperonnes d'histones forme un réseau complexe responsable du transport des histones, du facteur d'assemblage de la chromatine 1 et du maintien de l'intégrité du génome . Les réseaux de chaperonnes facilitent le transport des histones synthétisées dans le cytoplasme et qui doivent être escortées jusqu'au noyau cellulaire . Ce réseau garantit le dépôt des histones aux emplacements génomiques appropriés, préservant ainsi l'intégrité et la fonction de la chromatine .
Les chaperonnes d'histones jouent un rôle crucial dans la réponse aux lésions de l'ADN en régulant l'accessibilité de la chromatine. Par exemple, en réponse aux cassures double brin , des chaperonnes telles que FACT et ASF1 contribuent à la désassemblage des nucléosomes au niveau des lésions, permettant ainsi aux facteurs de réparation d'accéder à la lésion. Une fois la réparation terminée, ces chaperonnes facilitent le réassemblage des nucléosomes , restaurant la structure de la chromatine et assurant le maintien de l'information épigénétique.
Outre leur rôle dans la stabilité du génome, les chaperonnes d'histones contribuent à l'hérédité épigénétique . Lors de la division cellulaire, l'état de la chromatine doit être fidèlement transmis aux cellules filles. Les chaperonnes aident à répartir les histones parentales sur les brins d'ADN nouvellement synthétisés, préservant ainsi les modifications d'histones et assurant la continuité de l'identité cellulaire. Des perturbations dans ces processus peuvent entraîner des anomalies épigénétiques associées à des troubles du développement.
Synthèse
La première étape de la duplication de la structure de la chromatine est la synthèse des protéines histones : H1, H2A, H2B, H3 et H4. Ces protéines sont synthétisées durant la phase S du cycle cellulaire. Différents mécanismes contribuent à l’augmentation de la synthèse des histones.
Levure
Chez la levure, chaque gène d'histone possède une ou deux copies, dispersées sur les chromosomes et non regroupées. La transcription des gènes d'histones est contrôlée par de multiples protéines régulatrices, telles que des facteurs de transcription, qui se lient aux régions promotrices des histones. Chez la levure bourgeonnante, le gène candidat pour l'activation de l'expression des gènes d'histones est SBF. SBF est un facteur de transcription activé en fin de phase G1, lorsqu'il se dissocie de son répresseur Whi5 . Cette dissociation a lieu lorsque Whi5 est phosphorylé par Cdc8, une Cdk G1/S . La répression de l'expression des gènes d'histones en dehors des phases S dépend des protéines Hir, qui forment une structure de chromatine inactive au niveau des gènes d'histones, bloquant ainsi les activateurs de transcription
Métazoaire
Chez les métazoaires, l'augmentation du taux de synthèse des histones est due à une maturation accrue des pré-ARNm et à une diminution de la dégradation des ARNm, ce qui entraîne une augmentation de la quantité d'ARNm actif pour la traduction des protéines histones. Le mécanisme d'activation des ARNm repose sur l'élimination d'un segment de l'extrémité 3' du brin d'ARNm et dépend de l'association avec la protéine de liaison à la boucle tige ( SLBP ) . La SLBP stabilise également les ARNm des histones pendant la phase S en bloquant leur dégradation par la nucléase 3'hExo . Les niveaux de SLBP sont régulés par des protéines du cycle cellulaire, ce qui provoque leur accumulation lors de l'entrée en phase S et leur dégradation lors de la sortie de phase S. La SLBP est marquée pour la dégradation par phosphorylation de deux résidus thréonine par des kinases cycline-dépendantes, probablement la cycline A/cdk2, à la fin de la phase S. Les métazoaires possèdent également de multiples copies de gènes d'histones regroupées sur des chromosomes qui sont localisées dans des structures appelées corps de Cajal, comme déterminé par l'analyse de capture de conformation chromosomique à l'échelle du génome (4C-Seq).
Lien entre le contrôle du cycle cellulaire et la synthèse
La protéine nucléaire de l'ataxie-télangiectasie (NPAT), également connue sous le nom de coactivateur nucléaire de la transcription des histones, est un facteur de transcription qui active la transcription des gènes d'histones sur les chromosomes 1 et 6 des cellules humaines. NPAT est aussi un substrat de la cycline E-Cdk2, essentielle à la transition entre la phase G1 et la phase S. NPAT n'active l'expression des gènes d'histones qu'après avoir été phosphorylée par la cycline E-Cdk2, un complexe G1/S-Cdk, au début de la phase S . Ceci met en évidence un lien de régulation important entre le contrôle du cycle cellulaire et la synthèse des histones.
Histoire
Les histones ont été découvertes en 1884 par Albrecht Kossel . Le mot « histone » date de la fin du 19e siècle et est dérivé du mot allemand histon , un mot lui-même d'origine incertaine, peut-être du grec ancien ἵστημι (hístēmi, « faire tenir debout ») ou ἱστός (histós, « métier à tisser »).
Au début des années 1960, avant que les différents types d'histones ne soient identifiés et que leur forte conservation chez des organismes taxonomiquement divers ne soit établie, James F. Bonner et ses collaborateurs ont entrepris l'étude de ces protéines, connues pour être étroitement associées à l'ADN dans le noyau des organismes supérieurs. Bonner et son postdoctorant, Ru Chih C. Huang, ont démontré que la chromatine isolée ne permettait pas la transcription de l'ARN in vitro, mais que l'extraction des histones de la chromatine rendait possible la transcription de l'ARN à partir de l'ADN restant. Leur article est devenu une référence. En 1964, Paul T'so et James Bonner ont organisé un Congrès mondial sur la chimie et la biologie des histones, au cours duquel il est apparu clairement qu'aucun consensus n'existait quant au nombre de types d'histones et que l'on ignorait comment elles se compareraient une fois isolées de différents organismes. Bonner et ses collaborateurs ont ensuite mis au point des méthodes permettant de séparer chaque type d'histone, de purifier les histones individuellement, de comparer la composition en acides aminés d'une même histone chez différents organismes et de comparer les séquences d'acides aminés d'une même histone chez différents organismes, en collaboration avec Emil Smith de l'UCLA. Par exemple, ils ont constaté que la séquence de l'histone IV était hautement conservée entre le pois et le thymus de veau. Cependant, leurs travaux sur les caractéristiques biochimiques des histones individuelles n'ont pas permis de révéler comment les histones interagissaient entre elles ou avec l'ADN auquel elles étaient fortement liées.
Dans les années 1960 également, Vincent Allfrey et Alfred Mirsky avaient suggéré, sur la base de leurs analyses d'histones, que l'acétylation et la méthylation des histones pouvaient constituer un mécanisme de contrôle transcriptionnel. Cependant, ils ne disposaient pas d'analyses aussi détaillées que celles réalisées ultérieurement par d'autres chercheurs, permettant de démontrer la spécificité génique de cette régulation. Jusqu'au début des années 1990, la plupart des chercheurs considéraient les histones comme un matériau d'emballage inerte pour l'ADN nucléaire eucaryote. Cette conception reposait en partie sur les modèles de Mark Ptashne et d'autres, qui estimaient que la transcription était activée par des interactions protéine-ADN et protéine-protéine sur des matrices d'ADN largement nues, comme c'est le cas chez les bactéries.
Dans les années 1980, Yahli Lorch et Roger Kornberg ont montré qu'un nucléosome sur un promoteur central empêche l'initiation de la transcription in vitro, et Michael Grunstein a démontré que les histones répriment la transcription in vivo, ce qui a conduit à l'idée que le nucléosome est un répresseur génique général. La levée de cette répression impliquerait à la fois la modification des histones et l'action de complexes de remodelage de la chromatine. Vincent Allfrey et Alfred Mirsky avaient auparavant proposé un rôle de la modification des histones dans l'activation transcriptionnelle , considérée comme une manifestation moléculaire de l'épigénétique. Michael Grunstein et David Allis ont trouvé des arguments en faveur de cette proposition, notamment l'importance de l'acétylation des histones pour la transcription chez la levure et l'activité de l'activateur transcriptionnel Gcn5 en tant qu'histone acétyltransférase.
La découverte de l'histone H5 semble remonter aux années 1970, et elle est maintenant considérée comme une isoforme de l'histone H1 .