Un potentiel postsynaptique inhibiteur ( PPSI ) est un type de potentiel synaptique qui rend un neurone postsynaptique moins susceptible de générer un potentiel d'action . L'opposé d'un potentiel postsynaptique inhibiteur est un potentiel postsynaptique excitateur (PPSE), qui est un potentiel synaptique qui rend un neurone postsynaptique plus susceptible de générer un potentiel d'action. Les PPSE peuvent avoir lieu dans toutes les synapses chimiques, qui utilisent la sécrétion de neurotransmetteurs pour créer une signalisation de cellule à cellule. Les PPSE et les PPSE sont en compétition les uns avec les autres dans de nombreuses synapses d'un neurone. Cela détermine si un potentiel d'action se produisant au niveau du terminal présynaptique produit un potentiel d'action au niveau de la membrane postsynaptique. Certains neurotransmetteurs courants impliqués dans les PPSE sont le GABA et la glycine .
Les neurones présynaptiques inhibiteurs libèrent des neurotransmetteurs qui se lient ensuite aux récepteurs postsynaptiques ; cela induit un changement de la perméabilité de la membrane neuronale postsynaptique à des ions particuliers. Un courant électrique qui modifie le potentiel de la membrane postsynaptique pour créer un potentiel postsynaptique plus négatif est généré, c'est-à-dire que le potentiel de la membrane postsynaptique devient plus négatif que le potentiel de la membrane au repos, et c'est ce qu'on appelle l'hyperpolarisation . Pour générer un potentiel d'action, la membrane postsynaptique doit se dépolariser — le potentiel de la membrane doit atteindre un seuil de tension plus positif que le potentiel de la membrane au repos. Par conséquent, l'hyperpolarisation de la membrane postsynaptique réduit la probabilité que la dépolarisation se produise suffisamment pour générer un potentiel d'action dans le neurone postsynaptique.
La dépolarisation peut également se produire en raison d'un IPSP si le potentiel inverse se situe entre le seuil de repos et le seuil du potentiel d'action . Une autre façon d'envisager les potentiels postsynaptiques inhibiteurs est de considérer qu'ils constituent également un changement de conductance du chlorure dans la cellule neuronale, car ils diminuent la force motrice. En effet, si le neurotransmetteur libéré dans la fente synaptique provoque une augmentation de la perméabilité de la membrane postsynaptique aux ions chlorure en se liant aux canaux ioniques chlorure ligand-dépendants et en provoquant leur ouverture, alors les ions chlorure, qui sont en plus grande concentration dans la fente synaptique, diffusent dans le neurone postsynaptique. Comme il s'agit d'ions chargés négativement, une hyperpolarisation en résulte, ce qui réduit la probabilité qu'un potentiel d'action soit généré dans le neurone postsynaptique. Des microélectrodes peuvent être utilisées pour mesurer les potentiels postsynaptiques au niveau des synapses excitatrices ou inhibitrices.
En général, un potentiel postsynaptique dépend du type et de la combinaison du canal récepteur, du potentiel inverse du potentiel postsynaptique, du seuil de tension du potentiel d'action , de la perméabilité ionique du canal ionique, ainsi que des concentrations des ions à l'intérieur et à l'extérieur de la cellule ; cela détermine s'il est excitateur ou inhibiteur. Les IPSP ont toujours tendance à maintenir le potentiel membranaire plus négatif que le seuil du potentiel d'action et peuvent être considérés comme une « hyperpolarisation transitoire ».
Les IPSP ont été étudiés pour la première fois dans les motoneurones par David PC Lloyd, John Eccles et Rodolfo Llinás dans les années 1950 et 1960.

Composants
Types
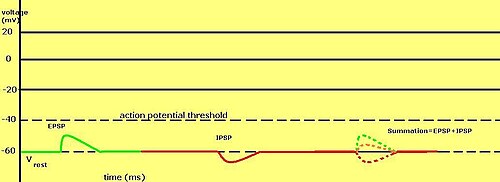
Ce système permet d'additionner temporellement les PPSE avec les PPSE sous-liminaires ou supraliminaires pour réduire l'amplitude du potentiel postsynaptique résultant. Les PPSE équivalents (positifs) et les PPSE (négatifs) peuvent s'annuler lorsqu'ils sont additionnés. L'équilibre entre les PPSE et les PPSE est très important dans l'intégration des informations électriques produites par les synapses inhibitrices et excitatrices.
Facteurs
La taille du neurone peut également affecter le potentiel postsynaptique inhibiteur. La simple sommation temporelle des potentiels postsynaptiques se produit dans les neurones plus petits, alors que dans les neurones plus grands, un plus grand nombre de synapses et de récepteurs ionotropes ainsi qu'une plus grande distance entre la synapse et le soma permettent la prolongation des interactions entre les neurones.
Neurotransmetteurs inhibiteurs
Le GABA est un neurotransmetteur très courant utilisé dans les IPSP dans le cerveau et la rétine des mammifères adultes. de glycine et leurs récepteurs fonctionnent de la même manière dans la moelle épinière, le cerveau et la rétine.
Récepteurs inhibiteurs
Il existe deux types de récepteurs inhibiteurs :
Récepteurs ionotropes
Les récepteurs ionotropes (également appelés canaux ioniques ligand-dépendants) jouent un rôle important dans les potentiels postsynaptiques inhibiteurs. Un neurotransmetteur se lie au site extracellulaire et ouvre le canal ionique constitué d'un domaine transmembranaire qui permet aux ions de traverser la membrane à l'intérieur de la cellule postsynaptique. Ce type de récepteur produit des actions postsynaptiques très rapides dans les quelques millisecondes suivant la réception d'un potentiel d'action par le terminal présynaptique. Ces canaux influencent l'amplitude et l'évolution temporelle des potentiels postsynaptiques dans leur ensemble.
Les récepteurs GABA ionotropes ( récepteurs GABA A ) sont des pentamères généralement composés de trois sous-unités différentes (α, β, γ), bien que plusieurs autres sous-unités (δ, ε, θ, π, ρ) et conformations existent. Les canaux ouverts sont sélectivement perméables aux ions chlorure ou potassium (selon le type de récepteur) et permettent à ces ions de traverser la membrane. Si le potentiel électrochimique de l'ion est plus négatif que celui du seuil du potentiel d'action, le changement de conductance résultant de la liaison du GABA à ses récepteurs maintient le potentiel postsynaptique plus négatif que le seuil et diminue la probabilité que le neurone postsynaptique termine un potentiel d'action.
Les récepteurs GABA ionotropes sont utilisés pour la liaison de divers médicaments tels que les barbituriques ( phénobarbital , pentobarbital ), les stéroïdes et la picrotoxine . Les benzodiazépines (Valium) se lient aux sous-unités α et γ des récepteurs GABA pour améliorer la signalisation GABAergique. L'alcool module également les récepteurs GABA ionotropes.
Récepteurs métabotropiques
Les récepteurs métabotropes sont souvent des récepteurs couplés à la protéine G tels que les récepteurs GABA B. Ceux-ci n'utilisent pas de canaux ioniques dans leur structure ; ils se composent plutôt d'un domaine extracellulaire qui se lie à un neurotransmetteur et d'un domaine intracellulaire qui se lie à la protéine G. Cela déclenche l'activation de la protéine G, qui se libère ensuite du récepteur et interagit avec les canaux ioniques et d'autres protéines pour ouvrir ou fermer les canaux ioniques par l'intermédiaire de messagers intracellulaires. Ils produisent des réponses postsynaptiques lentes (de quelques millisecondes à quelques minutes) et peuvent être activés en conjonction avec des récepteurs ionotropes pour créer des potentiels postsynaptiques rapides et lents au niveau d'une synapse particulière. Les récepteurs métabotropes GABA, hétérodimères des sous-unités R1 et R2, utilisent des canaux potassiques au lieu des canaux chlorure. Ils peuvent également bloquer les canaux ioniques calciques pour hyperpolariser les cellules postsynaptiques.
Importance
Les applications des potentiels postsynaptiques inhibiteurs sont nombreuses dans le monde réel. Les médicaments qui affectent les actions du neurotransmetteur peuvent traiter les troubles neurologiques et psychologiques grâce à différentes combinaisons de types de récepteurs, de protéines G et de canaux ioniques dans les neurones postsynaptiques.
Par exemple, des études sont en cours sur la désensibilisation et le trafic des récepteurs médiés par les récepteurs opioïdes dans le locus coeruleus du cerveau. Lorsqu’une concentration élevée d’agoniste est appliquée pendant une durée prolongée (quinze minutes ou plus), l’hyperpolarisation atteint un pic puis diminue. Ce phénomène est important car il s’agit d’un prélude à la tolérance : plus on a besoin d’opioïdes pour soulager la douleur, plus le patient y est tolérant. Ces études sont importantes car elles nous aident à mieux comprendre comment nous gérons la douleur et nos réponses à diverses substances qui aident à la traiter. En étudiant notre tolérance à la douleur, nous pouvons développer des médicaments plus efficaces pour le traitement de la douleur.
En outre, des recherches sont menées dans le domaine des neurones dopaminergiques de l'aire tegmentale ventrale, qui traite de la récompense, et de la substance noire, qui est impliquée dans le mouvement et la motivation. Les réponses métabotropiques se produisent dans les neurones dopaminergiques par le biais de la régulation de l'excitabilité des cellules. Les opioïdes inhibent la libération de GABA, ce qui diminue le degré d'inhibition et leur permet de s'activer spontanément. La morphine et les opioïdes sont liés aux potentiels postsynaptiques inhibiteurs car ils induisent une désinhibition dans les neurones dopaminergiques.
Les IPSP peuvent également être utilisés pour étudier les caractéristiques d'entrée-sortie d'une synapse inhibitrice du cerveau antérieur utilisée pour étudier plus en détail le comportement appris, par exemple dans une étude sur l'apprentissage du chant chez les oiseaux à l'Université de Washington. Des trains de Poisson d'IPSP unitaires ont été induits à une fréquence élevée pour reproduire les pics postsynaptiques dans la partie médiane du noyau thalamique dorso-latéral sans aucune entrée excitatrice supplémentaire. Cela montre un excès d'activation GABAergique thalamique. Ceci est important car le timing des pics est nécessaire pour une localisation correcte du son dans les voies auditives ascendantes. Les oiseaux chanteurs utilisent des terminaux synaptiques caliciels GABAergiques et une synapse de type calcyx de telle sorte que chaque cellule du noyau thalamique dorso-latéral reçoit au plus deux terminaux axonaux des noyaux gris centraux pour créer de grands courants postsynaptiques.
Les potentiels postsynaptiques inhibiteurs sont également utilisés pour étudier les noyaux gris centraux des amphibiens afin de voir comment la fonction motrice est modulée par ses sorties inhibitrices du striatum au tectum et au tegmentum. Les comportements guidés visuellement peuvent être régulés par la voie inhibitrice striato-tegmentale trouvée chez les amphibiens dans une étude réalisée au Baylor College of Medicine et à l'Académie chinoise des sciences. Les noyaux gris centraux des amphibiens sont très importants pour recevoir des entrées visuelles, auditives, olfactives et mécanosensorielles ; la voie désinhibitrice striato-protecto-tectale est importante dans les comportements de capture de proies des amphibiens. Lorsque le striatum ipsilatéral d'un crapaud adulte était stimulé électriquement, des potentiels postsynaptiques inhibiteurs étaient induits dans les neurones tegmentaux binoculaires, ce qui affectait le système visuel du crapaud.
Études
Les potentiels postsynaptiques inhibiteurs peuvent eux-mêmes être inhibés par un processus de signalisation appelé « suppression de l'inhibition induite par dépolarisation (DSI) » dans les cellules pyramidales CA1 et les cellules de Purkinje cérébelleuses. Dans un cadre de laboratoire, des dépolarisations par étapes du soma ont été utilisées pour créer des DSI, mais elles peuvent également être obtenues par une dépolarisation induite synaptiquement des dendrites. Les DSI peuvent être bloqués par des antagonistes des canaux calciques des récepteurs ionotropes sur les somas et les dendrites apicales proximales des cellules pyramidales CA1. Les potentiels postsynaptiques inhibiteurs dendritiques peuvent être sévèrement réduits par les DSI par dépolarisation directe.
Dans cette optique, les potentiels postsynaptiques inhibiteurs sont utiles dans la signalisation du bulbe olfactif au cortex olfactif . Les EPSP sont amplifiés par la conductance persistante des ions sodium dans les cellules touffues externes . La conductance des ions calcium activée à basse tension améliore encore les EPSP plus gros. La conductance cationique non sélective activée par hyperpolarisation diminue la somme et la durée des EPSP et transforme également les entrées inhibitrices en excitation postsynaptique. Les IPSP entrent en scène lorsque les membranes des cellules touffues sont dépolarisées et les IPSP provoquent alors une inhibition. Au seuil de repos, les IPSP induisent des potentiels d'action. Le GABA est responsable d'une grande partie du travail des IPSP dans les cellules touffues externes.
Une autre étude intéressante des potentiels postsynaptiques inhibiteurs porte sur les oscillations du rythme thêta neuronal qui peuvent être utilisées pour représenter des phénomènes électrophysiologiques et divers comportements. Les rythmes thêta se trouvent dans l' hippocampe et l'inhibition synaptique GABAergique permet de les moduler. Ils dépendent des IPSP et sont déclenchés soit dans le CA3 par des récepteurs muscariniques de l'acétylcholine, soit dans le C1 par l'activation des récepteurs métabotropiques du glutamate du groupe I. Lorsque les interneurones sont activés par des récepteurs métabotropiques de l'acétylcholine dans la région CA1 des tranches d'hippocampe de rat, un modèle thêta d'IPSP dans les cellules pyramidales se produit indépendamment de l'entrée. Cette recherche étudie également les DSI, montrant que les DSI interrompent le rythme initié par l'acétylcholine métabotropique par la libération d'endocannabinoïdes. Un mécanisme dépendant des endocannabinoïdes peut perturber les IPSP thêta par des potentiels d'action délivrés sous forme de modèle d'éclatement ou de train bref. De plus, l’activation des récepteurs métabotropiques du glutamate supprime toute activité IPSP thêta via une voie indépendante de la protéine G et des ions calcium.
Les potentiels postsynaptiques inhibiteurs ont également été étudiés dans la cellule de Purkinje par amplification dendritique. L'étude s'est concentrée sur la propagation des IPSP le long des dendrites et sa dépendance aux récepteurs ionotropes en mesurant l'amplitude et l'évolution temporelle du potentiel postsynaptique inhibiteur. Les résultats ont montré que les potentiels postsynaptiques inhibiteurs composés et unitaires sont amplifiés par les canaux calciques dendritiques. La largeur d'un IPSP somatique est indépendante de la distance entre le soma et la synapse alors que le temps de montée augmente avec cette distance. Ces IPSP régulent également les rythmes thêta dans les cellules pyramidales. D'autre part, les potentiels postsynaptiques inhibiteurs sont dépolarisants et parfois excitateurs dans les neurones spinaux immatures des mammifères en raison des concentrations élevées de chlorure intracellulaire via les canaux ionotropes GABA ou chlorure de glycine. Ces dépolarisations activent les canaux calciques voltage-dépendants. Ils deviennent plus tard hyperpolarisants à mesure que le mammifère mûrit. Plus précisément, chez les rats, cette maturation se produit pendant la période périnatale lorsque les projections du tronc cérébral atteignent l'élargissement lombaire. Des entrées modulatrices descendantes sont nécessaires au passage développemental des potentiels postsynaptiques inhibiteurs dépolarisants aux potentiels postsynaptiques inhibiteurs hyperpolarisants. Cela a été étudié par des transections complètes de la moelle épinière à la naissance des rats et l'enregistrement des IPSP des motoneurones lombaires à la fin de la première semaine après la naissance.
Le glutamate , un neurotransmetteur excitateur, est généralement associé à des potentiels postsynaptiques excitateurs dans la transmission synaptique. Cependant, une étude réalisée au Vollum Institute de l'Oregon Health Sciences University démontre que le glutamate peut également être utilisé pour induire des potentiels postsynaptiques inhibiteurs dans les neurones. Cette étude explique que les récepteurs métabotropiques du glutamate présentent des protéines G activées dans les neurones dopaminergiques qui induisent l'hydrolyse des phosphoinositides. Les produits résultants se lient aux récepteurs d'inositol triphosphate (IP3) par l'intermédiaire des canaux ioniques calciques. Le calcium provient des réserves et active la conductance potassique, ce qui provoque une inhibition pure dans les cellules dopaminergiques. Les niveaux changeants de glutamate libéré par voie synaptique créent une excitation par l'activation des récepteurs ionotropiques, suivie de l'inhibition des récepteurs métabotropiques du glutamate.