.svg/1280px-Metapopulation_(1).svg.png)
Une métapopulation est un groupe de populations spatialement séparées de la même espèce qui interagissent à un certain niveau. Le terme métapopulation a été inventé par Richard Levins en 1969 pour décrire un modèle de dynamique de population d'insectes nuisibles dans les champs agricoles, mais l'idée a été plus largement appliquée aux espèces vivant dans des habitats naturellement ou artificiellement fragmentés . Selon les propres termes de Levins, il s'agit d'une « population de populations ».
On considère généralement qu'une métapopulation est constituée de plusieurs populations distinctes ainsi que de zones d'habitat adaptées actuellement inoccupées. Dans la théorie classique des métapopulations, chaque population évolue de manière relativement indépendante des autres populations et finit par disparaître en raison de la stochasticité démographique (fluctuations de la taille de la population dues à des événements démographiques aléatoires ) ; plus la population est petite, plus les risques de dépression de consanguinité et d'extinction sont élevés.
Bien que les populations individuelles aient une durée de vie limitée, la métapopulation dans son ensemble est souvent stable car les immigrants d'une population (qui peut, par exemple, connaître un boom démographique) sont susceptibles de recoloniser un habitat laissé libre par l'extinction d'une autre population. Ils peuvent également émigrer vers une petite population et sauver cette population de l'extinction (ce que l'on appelle l' effet de sauvetage ). Un tel effet de sauvetage peut se produire parce que les populations en déclin laissent des opportunités de niche ouvertes aux « sauveteurs ».
Le développement de la théorie des métapopulations, en conjonction avec le développement de la dynamique source-puits , a souligné l’importance de la connectivité entre des populations apparemment isolées. Bien qu’aucune population ne puisse à elle seule garantir la survie à long terme d’une espèce donnée, l’effet combiné de plusieurs populations peut y parvenir.
La théorie de la métapopulation a d'abord été développée pour les écosystèmes terrestres, puis appliquée au monde marin. En sciences halieutiques, le terme « sous-population » est équivalent au terme scientifique de métapopulation « population locale ». La plupart des exemples marins sont fournis par des espèces relativement sédentaires occupant des parcelles d'habitat distinctes , avec à la fois un recrutement local et un recrutement provenant d'autres populations locales dans la métapopulation plus large. Kritzer et Sale ont plaidé contre l'application stricte des critères de définition de la métapopulation selon lesquels les risques d'extinction des populations locales doivent être non négligeables.
Le biologiste finlandais Ilkka Hanski de l’ Université d’Helsinki a été un contributeur important à la théorie des métapopulations.
Prédation et oscillations
Les premières expériences sur la prédation et l'hétérogénéité spatiale ont été menées par GF Gause dans les années 1930, sur la base de l' équation Lotka-Volterra , formulée au milieu des années 1920, mais aucune autre application n'a été réalisée. L'équation Lotka-Volterra suggérait que la relation entre les prédateurs et leurs proies entraînerait des oscillations de population au fil du temps en fonction des densités initiales de prédateurs et de proies. Les premières expériences de Gause pour prouver les oscillations prédites par cette théorie ont échoué parce que les interactions prédateur-proie n'étaient pas influencées par l'immigration. Cependant, une fois l'immigration introduite, les cycles de population ont décrit avec précision les oscillations prédites par l'équation Lotka-Volterra, avec les pics d'abondance des proies légèrement décalés vers la gauche des pics des densités de prédateurs. Les expériences de Huffaker ont étendu celles de Gause en examinant comment les facteurs de migration et d'hétérogénéité spatiale conduisent à des oscillations prédateur-proie.
Expériences de Huffaker sur les interactions prédateur-proie (1958)
Afin d'étudier la prédation et les oscillations de population, Huffaker a utilisé des espèces d'acariens, l'une étant le prédateur et l'autre la proie. Il a mis en place une expérience contrôlée en utilisant des oranges, dont la proie se nourrissait, comme habitat spatialement structuré dans lequel le prédateur et la proie interagiraient. Au début, Huffaker a rencontré des difficultés similaires à celles de Gause pour créer une interaction prédateur-proie stable. En utilisant uniquement des oranges, les espèces proies ont rapidement disparu, suivies par conséquent de l'extinction des prédateurs. Cependant, il a découvert qu'en modifiant la structure spatiale de l'habitat, il pouvait manipuler la dynamique de population et permettre au taux de survie global des deux espèces d'augmenter. Il y est parvenu en modifiant la distance entre la proie et les oranges (leur nourriture), en établissant des barrières au mouvement des prédateurs et en créant des couloirs pour que la proie se disperse. Ces changements ont entraîné une augmentation des parcelles d'habitat et, à leur tour, ont fourni davantage de zones où la proie peut rechercher une protection temporaire. Lorsque les proies disparaissaient localement dans une parcelle d'habitat, elles pouvaient se rétablir en migrant vers de nouvelles parcelles avant d'être attaquées par des prédateurs. Cette structure spatiale de l'habitat par parcelles permettait la coexistence entre les espèces prédatrices et proies et favorisait un modèle d'oscillation de population stable. Bien que le terme métapopulation n'ait pas encore été inventé, les facteurs environnementaux d' hétérogénéité spatiale et de répartition de l'habitat décriraient plus tard les conditions d'une métapopulation liées à la façon dont des groupes de populations d'espèces spatialement séparées interagissent les unes avec les autres. L'expérience de Huffaker est importante car elle a montré comment les métapopulations peuvent affecter directement les interactions prédateur-proie et, à leur tour, influencer la dynamique de population.
Le modèle de Levin
Le modèle original de Levins s'appliquait à une métapopulation répartie sur de nombreuses parcelles d'habitat propice avec beaucoup moins d'interactions entre les parcelles qu'au sein d'une même parcelle. La dynamique de population au sein d'une parcelle a été simplifiée au point que seules la présence et l'absence ont été prises en compte. Chaque parcelle de son modèle est soit peuplée, soit non peuplée.
Soit N la fraction des parcelles occupées à un instant donné. Pendant un instant dt , chaque parcelle occupée peut devenir inoccupée avec une probabilité d'extinction edt . De plus, 1 − N parcelles sont inoccupées. En supposant un taux constant c de génération de propagules à partir de chacune des N parcelles occupées, pendant un instant dt , chaque parcelle inoccupée peut devenir occupée avec une probabilité de colonisation cNdt . En conséquence, le taux de changement temporel des parcelles occupées, dN/dt , est
Cette équation est mathématiquement équivalente au modèle logistique , avec une capacité de charge K donnée par
et le taux de croissance r
À l’équilibre, une certaine fraction de l’habitat de l’espèce sera donc toujours inoccupée.
Stochasticité et métapopulations
Les études de Huffaker , des modèles intégrant des facteurs stochastiques ont été créés. Ces modèles ont montré que la combinaison de la variabilité environnementale (stochasticité) et de taux de migration relativement faibles provoque une persistance indéfinie ou imprévisible. Cependant, l'expérience de Huffaker garantissait presque une persistance infinie en raison de la variable d'immigration contrôlée.
Modèles d'occupation de patchs stochastiques (SPOM)
L'un des principaux inconvénients du modèle de Levins est qu'il est déterministe , alors que les processus fondamentaux de métapopulation sont stochastiques . Les métapopulations sont particulièrement utiles pour discuter des espèces dans des habitats perturbés et de la viabilité de leurs populations , c'est-à-dire de leur probabilité d' extinction dans un intervalle de temps donné. Le modèle de Levins ne peut pas résoudre ce problème. Une façon simple d'étendre le modèle de Levins pour y intégrer des considérations spatiales et stochastiques consiste à utiliser le processus de contact . De simples modifications de ce modèle peuvent également être intégrées à la dynamique des parcelles . À un seuil de percolation donné , les effets de fragmentation de l'habitat se produisent dans ces configurations, prédisant des seuils d'extinction plus drastiques.
Pour des raisons de biologie de la conservation , les modèles de métapopulation doivent inclure (a) la nature finie des métapopulations (combien de parcelles sont propices à l'habitat) et (b) la nature probabiliste de l'extinction et de la colonisation. De plus, pour pouvoir appliquer ces modèles, les extinctions et les colonisations des parcelles doivent être asynchrones.
Microhabitats et métapopulations bactériennes
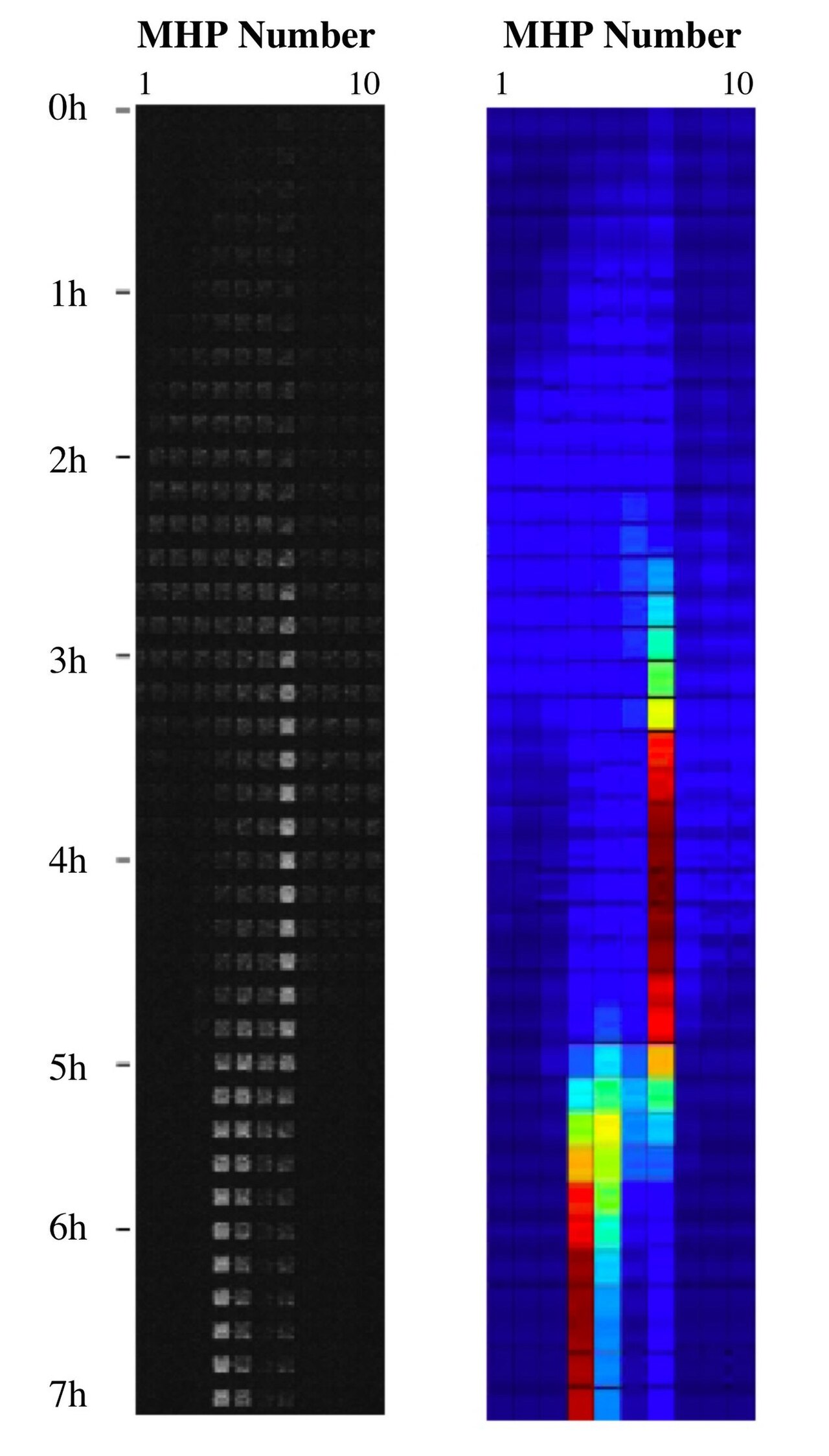
En combinant la nanotechnologie avec l'écologie du paysage , des paysages d'habitats synthétiques ont été fabriqués sur une puce en construisant une collection de mini-habitats bactériens avec des canaux à l'échelle nanométrique leur fournissant des nutriments pour le renouvellement de l'habitat et en les reliant par des couloirs dans différentes dispositions topologiques, générant une mosaïque spatiale de parcelles d'opportunités distribuées dans le temps. Cela peut être utilisé pour des expériences de paysage en étudiant les métapopulations bactériennes sur la puce, par exemple leur écologie évolutive .
Évolution du cycle de vie
Des modèles de métapopulation ont été utilisés pour expliquer l'évolution du cycle biologique, comme la stabilité écologique de la métamorphose des amphibiens dans les petits étangs printaniers . Des stratégies écologiques alternatives ont été développées. Par exemple, certaines salamandres renoncent à la métamorphose et atteignent la maturité sexuelle en tant que néotènes aquatiques . La durée saisonnière des zones humides et l'aire de migration des espèces déterminent quels étangs sont connectés et s'ils forment une métapopulation. La durée des stades du cycle biologique des amphibiens par rapport à la durée de l'étang printanier avant qu'il ne s'assèche régule le développement écologique des métapopulations reliant les parcelles aquatiques aux parcelles terrestres.