Le complexe microprocesseur est un complexe protéique impliqué dans les premières étapes de la maturation des microARN (miARN) et de l'interférence ARN (ARNi) dans les cellules animales. Ce complexe est composé au minimum de l' enzyme ribonucléase Drosha et de la protéine dimérique de liaison à l'ARN DGCR8 (également appelée Pasha chez les animaux non humains), et clive les substrats primaires des miARN en pré-miARN dans le noyau cellulaire . Le complexe microprocesseur est également le plus petit des deux complexes multiprotéiques contenant la protéine Drosha humaine .
Composition
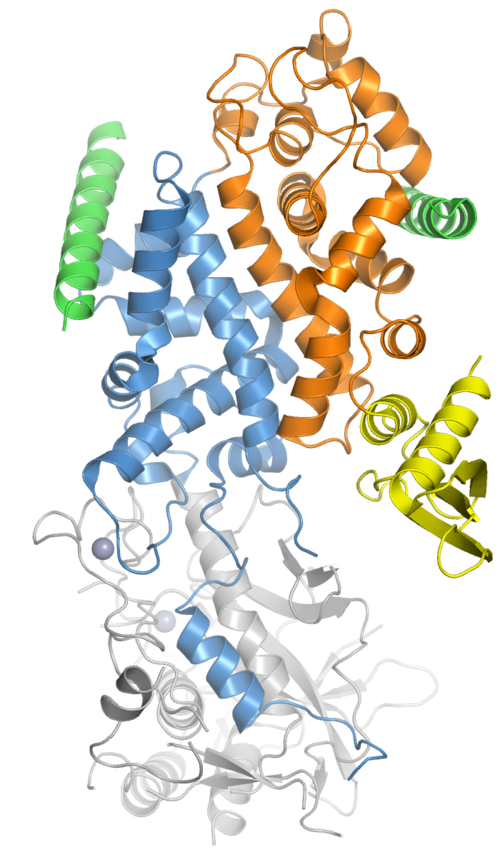
Le complexe microprocesseur est constitué au minimum de deux protéines : Drosha , une ribonucléase III ; et DGCR8 , une protéine de liaison à l’ARN double brin . (DGCR8 est le nom utilisé en génétique des mammifères, abréviation de « DiGeorge syndrome critical region 8 » ; la protéine homologue chez les organismes modèles tels que les mouches et les vers est appelée Pasha , pour « Partner of Drosha » .) La stœchiométrie du complexe minimal était autrefois difficile à déterminer expérimentalement, mais il a été démontré qu’il s’agit d’un hétérotrimère composé de deux protéines DGCR8 et d’une protéine Drosha.
Outre les composants catalytiquement actifs du microprocesseur, d'autres cofacteurs tels que les hélicases à ARN de type DEAD box et les ribonucléoprotéines nucléaires hétérogènes peuvent être présents dans le complexe pour moduler l'activité de Drosha . Certains miARN ne sont traités par le microprocesseur qu'en présence de cofacteurs spécifiques.
Fonction
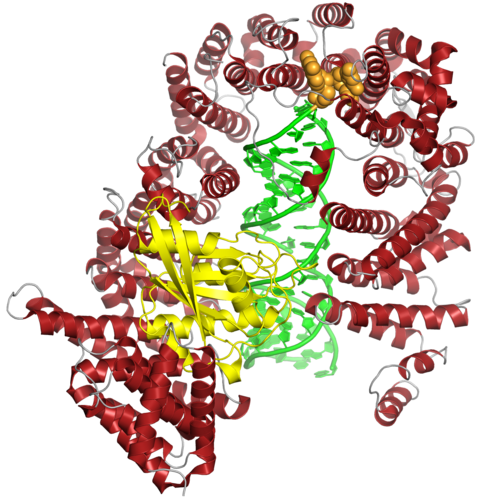
Situé dans le noyau cellulaire , le complexe microprocesseur clive les microARN primaires (pri-miARN) en précurseurs de microARN (pré-miARN) . Ses deux sous-unités sont nécessaires et suffisantes pour la maturation des pri-miARN en microARN . Ces molécules d'environ 70 nucléotides possèdent une structure en tige-boucle ou en épingle à cheveux. Les substrats pri-miARN peuvent provenir de gènes d'ARN non codants ou d' introns . Dans ce dernier cas, il a été démontré que le complexe microprocesseur interagit avec le spliceosome et que la maturation pri-miARN a lieu avant l'épissage .
Le clivage des pri-miARN par le microprocesseur se produit généralement de manière co- transcriptionnelle et laisse un surplomb simple brin caractéristique de 2 à 3 nucléotides, caractéristique de la RNase III, qui sert d'élément de reconnaissance pour la protéine de transport exportine-5 . Les pré-miARN sont exportés du noyau vers le cytoplasme de manière dépendante de RanGTP et sont ensuite traités, généralement par l' enzyme endoribonucléase Dicer .
L'hémine permet une maturation accrue des pri-miARN grâce à un changement conformationnel induit de la sous-unité DGCR8, et renforce également la spécificité de liaison de DGCR8 à l'ARN. DGCR8 reconnaît les jonctions entre les structures en épingle à cheveux et l'ARN simple brin et oriente Drosha pour cliver à environ 11 nucléotides de ces jonctions. Après le clivage et la dissociation de Drosha, DGCR8 reste en contact avec les pri-miARN.
Bien que la grande majorité des microARN (miARN) soient traités par le complexe microprocesseur, quelques exceptions, appelées mirtrons, ont été décrites ; il s’agit de très petits introns qui, après épissage, possèdent la taille et la structure en tige-boucle appropriées pour servir de pré-miARN. Les voies de maturation des miARN et des petits ARN interférents exogènes convergent au niveau du clivage par Dicer et sont largement identiques en aval. De manière générale, ces deux voies constituent l’interférence ARN (ARNi) . Le complexe microprocesseur est également impliqué dans la biogenèse ribosomique, notamment dans l’élimination des boucles R et l’activation de la transcription des gènes codant pour les protéines ribosomiques.
Règlement
La régulation génique par les microARN est largement répandue dans de nombreux génomes ; selon certaines estimations, plus de 60 % des gènes codant pour des protéines humaines seraient régulés par des microARN , bien que la qualité des preuves expérimentales concernant les interactions microARN-cible soit souvent faible . Étant donné que le traitement par le microprocesseur est un déterminant majeur de l’abondance des microARN, le microprocesseur lui-même constitue une cible importante de régulation.
Les protéines Drosha et DGCR8 sont toutes deux soumises à une régulation par modifications post-traductionnelles qui modulent leur stabilité, leur localisation intracellulaire et leur niveau d'activité. L'activité contre des substrats spécifiques peut être régulée par des cofacteurs protéiques supplémentaires interagissant avec le complexe microprocesseur. La région en boucle de la structure en tige-boucle du pri-miARN constitue également un élément de reconnaissance pour les protéines régulatrices, qui peuvent activer ou inhiber la maturation des miARN cibles par le complexe microprocesseur.
Le microprocesseur est lui-même autorégulé par rétroaction négative via son association avec une structure en épingle à cheveux de type pri-miRNA présente dans l' ARNm de DGCR8 , dont le clivage réduit l'expression de DGCR8 . Dans ce cas, la structure est située dans un exon et il est peu probable qu'elle fonctionne elle-même comme un miRNA.
Évolution
Drosha présente une ressemblance structurale frappante avec la ribonucléase Dicer , située en aval, suggérant une relation évolutive. Cependant, Drosha et les enzymes apparentées se trouvent uniquement chez les animaux, tandis que les homologues de Dicer sont largement distribués, y compris chez les protozoaires . Les deux composants du complexe microprocesseur sont conservés chez la grande majorité des métazoaires dont le génome est connu. Mnemiopsis leidyi , un cténophore , est dépourvu d'homologues de Drosha et de DGCR8 , ainsi que de microARN identifiables, et constitue le seul métazoaire connu pour lequel aucune trace génomique de Drosha n'a été détectée . Chez les plantes, la voie de biogenèse des microARN est quelque peu différente ; ni Drosha ni DGCR8 ne possèdent d' homologue dans les cellules végétales, où la première étape de la maturation des microARN est généralement réalisée par une autre ribonucléase nucléaire , DCL1 , homologue de Dicer .
Des analyses phylogénétiques suggèrent que les composants clés de l'interférence ARN basée sur des substrats exogènes étaient présents chez l' eucaryote ancestral , probablement comme mécanisme immunitaire contre les virus et les éléments transposables . L'élaboration de cette voie de régulation génique par les microARN serait apparue plus tard.
Signification clinique
L'implication des microARN dans les maladies a conduit les scientifiques à s'intéresser davantage au rôle de complexes protéiques supplémentaires, comme le complexe microprocesseur, capables d'influencer ou de moduler la fonction et l'expression des microARN. Le composant DGCR8 du complexe microprocesseur est affecté par la microdélétion 22q11.2 , une petite portion du chromosome 22. Cette délétion entraîne un traitement anormal des microARN , ce qui conduit au syndrome de DiGeorge .