"

En biologie évolutive du développement , l'hétérochronie désigne toute différence génétiquement contrôlée dans le calendrier, le rythme ou la durée d'un processus de développement chez un organisme , comparée à celle de ses ancêtres ou d'autres organismes. Ceci entraîne des modifications de la taille, de la forme, des caractéristiques, voire de la présence de certains organes et structures. Elle s'oppose à l' hétérotopie , un changement de position spatiale d'un processus au sein de l'embryon, qui peut également engendrer une innovation morphologique . L'hétérochronie se divise en hétérochronie intraspécifique, variation au sein d'une même espèce, et en hétérochronie interspécifique, variation phylogénétique , c'est-à-dire variation d'une espèce descendante par rapport à une espèce ancestrale.
Ces modifications influent toutes sur le début, la fin, le rythme ou la durée d'un processus de développement donné. Le concept d'hétérochronie a été introduit par Ernst Haeckel en 1875 et son sens moderne a été donné par Gavin de Beer en 1930.
En 1928, l’embryologiste anglais Walter Garstang a démontré que les larves de tuniciers partageaient des structures telles que la notochorde avec les vertébrés adultes , et a suggéré que les vertébrés étaient issus d’ une pédomorphose (néoténie) à partir de telles larves. Cette proposition impliquait (si elle était correcte) une phylogénie commune aux tuniciers et aux vertébrés, et que l’hétérochronie constituait un mécanisme principal de l’évolution.
La biologie évolutive du développement (evo-devo) moderne étudie la génétique moléculaire du développement. Elle cherche à expliquer chaque étape de la formation d'un organisme adulte à partir d'un zygote indifférencié en termes de contrôle de l' expression d'un gène après l'autre. De plus, elle relie ces mécanismes de contrôle du développement à la phylogénie . De Beer a, dans une certaine mesure, anticipé cette science de la fin du XXe siècle dans son ouvrage de 1930, Embryos and Ancestors démontrant que l'évolution pouvait se produire par hétérochronie, comme dans le cas de la pédomorphose, c'est-à-dire la conservation de caractères juvéniles chez l'adulte . De Beer soutenait que cela permettait des changements évolutifs rapides, trop brefs pour être enregistrés dans les archives fossiles , expliquant ainsi la probabilité de lacunes apparentes
Mécanismes
L’hétérochronie interspécifique désigne les différences de rythme ou de chronologie d’une espèce descendante par rapport à son ancêtre. Cela peut se traduire par une pédomorphose (troncation de l’ontogenèse ancestrale), une péramorphose (prolongement au-delà de l’ontogenèse ancestrale) ou une isomorphose (atteinte du même état ancestral par un mécanisme différent).
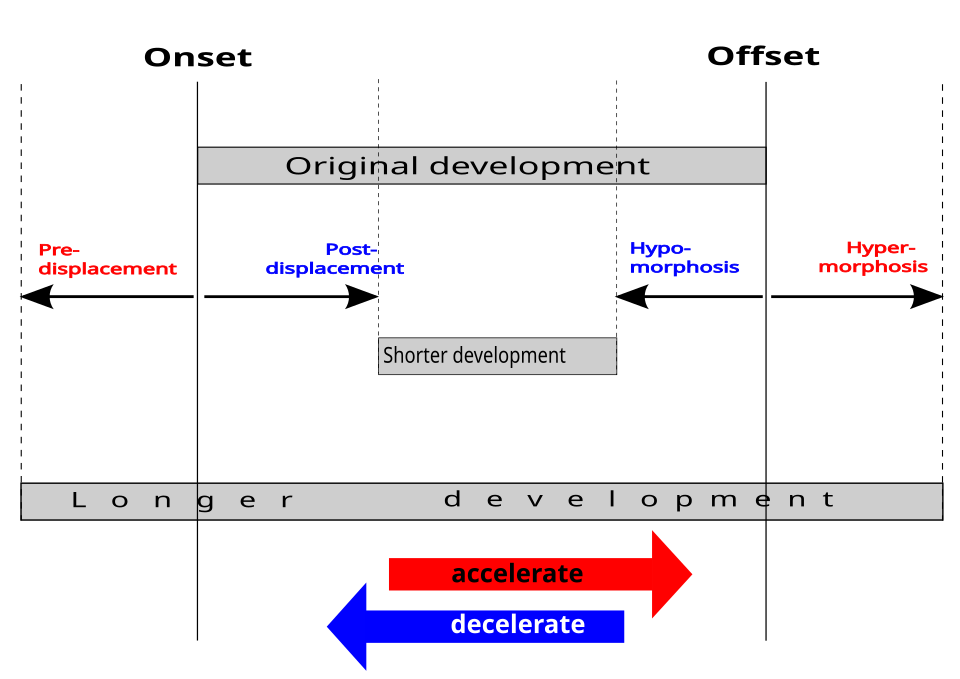
Il existe trois mécanismes majeurs d'hétérochronie chacun pouvant évoluer dans deux directions, donnant lieu à six types de perturbations, combinables de diverses manières . Ces perturbations entraînent un développement prolongé, décalé ou tronqué d'un processus particulier, comme l'action d'un gène codant pour un seul gène , par rapport à l'état ancestral ou à d'autres individus de la même espèce, selon que l'hétérochronie soit interspécifique ou intraspécifique. Identifier la perturbation en cause est crucial pour comprendre le mécanisme sous-jacent à la péramorphose ou à la pédomorphose
- Début : Un processus de développement peut commencer soit plus tôt, avant le déplacement , prolongeant ainsi son développement, soit plus tard, après le déplacement , le tronquant.
- Décalage : Un processus peut se terminer soit plus tard, par hypermorphose ( prolongement de son développement), soit plus tôt, par hypomorphose ou progénèse (troncation de celui-ci).
- Vitesse : La vitesse d'un processus peut s'accélérer, prolongeant son développement, ou ralentir (comme dans la néoténie), le tronquant.
Un exemple frappant de la façon dont l'accélération peut modifier l' organisation corporelle est celui des serpents . Alors qu'un vertébré typique comme la souris ne possède qu'une soixantaine de vertèbres, les serpents en ont entre 150 et 400, ce qui leur confère une colonne vertébrale extrêmement longue et leur permet une locomotion sinueuse . Les embryons de serpent y parviennent en accélérant leur système de formation des somites (segments du corps), qui repose sur un oscillateur. L'horloge de cet oscillateur fonctionne environ quatre fois plus vite chez les embryons de serpent que chez ceux de souris, créant initialement des somites très fins. Ceux-ci s'étendent ensuite pour adopter une forme typique des vertébrés, allongeant ainsi le corps.
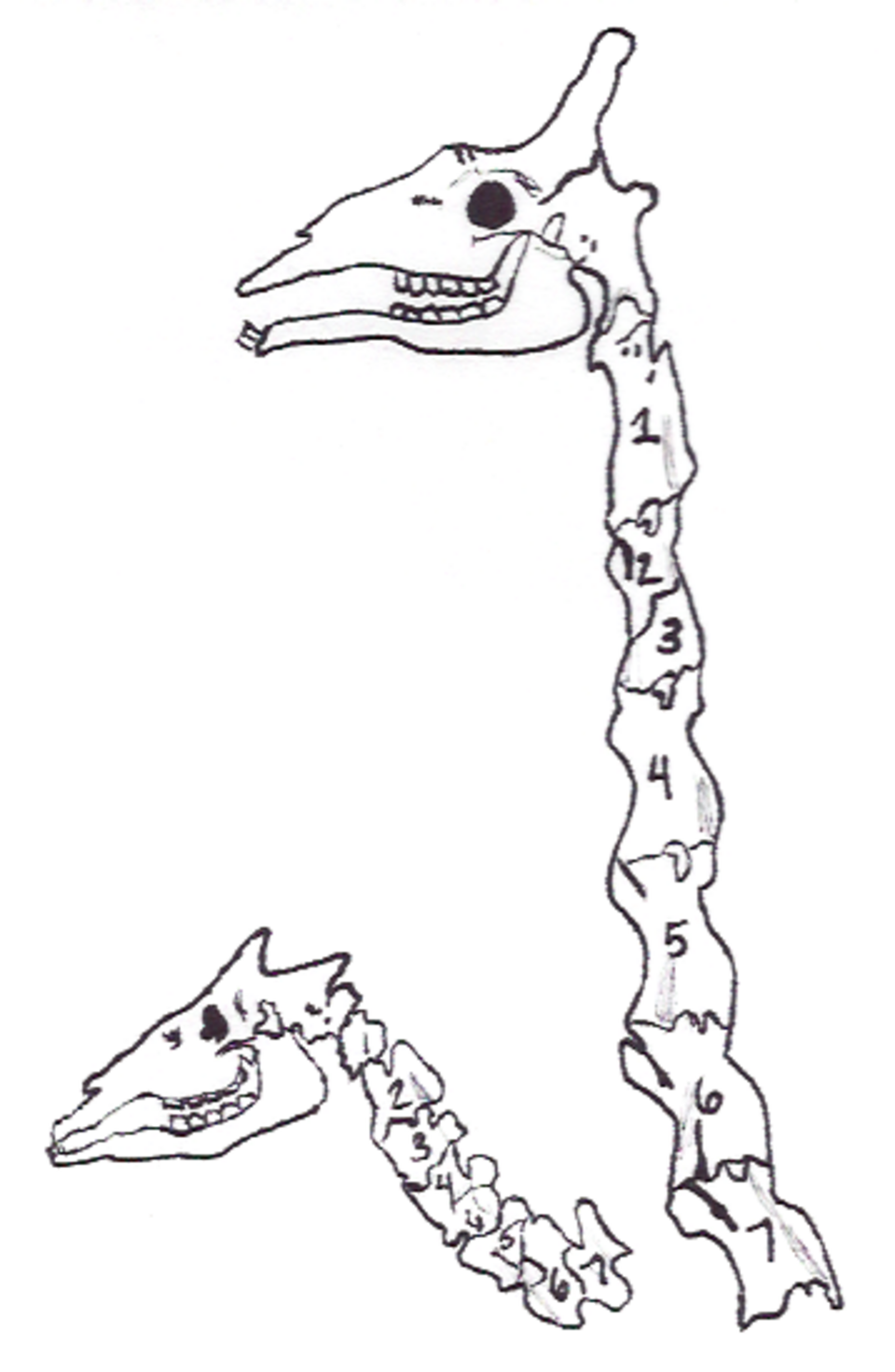
Les girafes acquièrent leur long cou grâce à une hétérochronie différente, prolongeant le développement de leurs vertèbres cervicales ; elles conservent le nombre habituel de ces vertèbres chez les mammifères, soit sept. Ce nombre semble être limité par l’utilisation des somites du cou pour former le diaphragme des mammifères ; de ce fait, le cou embryonnaire est divisé en trois modules, le module médian (C3 à C5) servant au diaphragme. On suppose que perturber ce processus entraînerait la mort de l’embryon plutôt que de lui donner davantage de vertèbres.
Détection
L'hétérochronie peut être identifiée en comparant des espèces phylogénétiquement proches, par exemple un groupe d'espèces d'oiseaux différentes dont les pattes ont des longueurs moyennes différentes. Ces comparaisons sont complexes car il n'existe pas de marqueurs temporels ontogénétiques universels. La méthode d'appariement d'événements tente de pallier cette difficulté en comparant la chronologie relative de deux événements à la fois . Cette méthode détecte les hétérochronies d'événements, par opposition aux changements allométriques . Son utilisation est fastidieuse car le nombre de caractères de paires d'événements augmente avec le carré du nombre d'événements comparés. L'appariement d'événements peut cependant être automatisé, par exemple avec le script PARSIMOV . Une méthode récente, l'analyse continue, repose sur une simple standardisation du temps ontogénétique ou des séquences, sur la parcimonie des changements au carré et sur les contrastes phylogénétiques indépendants .
Effets
Pédomorphose
La pédomorphose peut résulter d'une néoténie , c'est-à-dire la persistance de caractères juvéniles à l'âge adulte suite à un retard de développement somatique, ou d'une progénèse, c'est-à-dire une accélération du développement permettant à la forme juvénile d'atteindre la maturité sexuelle. Cela signifie que lors d'une progénèse, la croissance des cellules germinales est accélérée par rapport à la normale ou à la néoténie ; tandis que la croissance des cellules somatiques est normale lors d'une progénèse, mais ralentie lors d'une néoténie.
La néoténie retarde le développement de l'organisme vers l'âge adulte et a été décrite comme une « enfance éternelle » . Dans cette forme d'hétérochronie, la période de développement proprement dite de l'enfance est prolongée, et certains processus développementaux qui se produisent normalement uniquement pendant l'enfance (comme l'accélération de la croissance cérébrale chez l'humain ) se prolongent également durant cette période. La néoténie a été impliquée comme cause développementale de plusieurs changements comportementaux, en raison d'une plasticité cérébrale accrue et d'une enfance prolongée
La progénèse (ou pédogenèse) s'observe chez l' axolotl ( Ambystoma mexicanum ). Les axolotls atteignent leur pleine maturité sexuelle tout en conservant leurs nageoires et leurs branchies (autrement dit, ils restent sous la forme juvénile de leurs ancêtres). Ils demeurent en milieu aquatique sous cette forme de développement tronquée, au lieu de gagner la terre ferme comme les autres espèces de salamandres sexuellement matures. Il s'agirait d'une forme d'hypomorphose (arrêt précoce du développement) à la fois hormonale et génétique . La métamorphose complète permettant à la salamandre d'accéder à la forme adulte est ainsi bloquée par ces deux facteurs
La pédomorphose par progénèse pourrait jouer un rôle crucial dans l'évolution crânienne des oiseaux . Le crâne et le bec des oiseaux adultes actuels conservent l'anatomie des dinosaures théropodes juvéniles dont ils descendent. Les oiseaux actuels possèdent des yeux et un cerveau volumineux par rapport au reste du crâne ; une caractéristique observée chez les oiseaux adultes qui représente (en gros) le stade juvénile d'un dinosaure. Un ancêtre aviaire juvénile (comme le Coelophysis ) aurait un museau court, de grands yeux, un palais fin, un os jugal étroit, des os postorbitaires hauts et fins, des adducteurs limités et une boîte crânienne courte et bulbeuse. En vieillissant, un tel organisme subirait d'importantes modifications de sa morphologie crânienne pour développer un crâne robuste aux os plus grands et imbriqués. Les oiseaux, cependant, conservent cette morphologie juvénile. Des expériences moléculaires suggèrent que le facteur de croissance des fibroblastes 8 (FGF8) et des membres de la voie de signalisation WNT ont favorisé la pédomorphose chez les oiseaux. Ces voies de signalisation sont connues pour jouer un rôle dans la morphogenèse faciale chez d'autres espèces de vertébrés. La conservation de l'état ancestral juvénile a induit d'autres modifications anatomiques, aboutissant à un crâne léger et très mobile, composé de nombreux petits os non superposés . On pense que cela a facilité l'évolution de la cinétique crânienne chez les oiseaux , qui a joué un rôle crucial dans leur succès écologique.
Péramorphose

Un autre exemple de péramorphose s'observe chez les rongeurs insulaires. Leurs caractéristiques comprennent le gigantisme, des joues et des dents plus larges, une portée réduite et une durée de vie plus longue. Leurs congénères vivant en milieu continental sont beaucoup plus petits. Les rongeurs insulaires ont développé ces caractéristiques pour s'adapter à l'abondance de nourriture et de ressources présentes sur leurs îles. Ces facteurs font partie d'un phénomène complexe appelé syndrome insulaire ou règle de Foster .
La salamandre fouisseuse , proche parente de l'axolotl, présente à la fois une pédomorphose et une péramorphose. La larve peut se développer dans les deux sens. La densité de population, la nourriture et la quantité d'eau peuvent influencer l'expression de l'hétérochronie. Une étude menée sur la salamandre fouisseuse en 1987 a mis en évidence qu'un pourcentage plus élevé d'individus devenaient pédomorphes en cas de faible densité de population larvaire dans un niveau d'eau constant, comparativement à une forte densité de population larvaire dans un milieu aquatique en voie d'assèchement . Ce résultat a conduit à l'hypothèse que les pressions sélectives exercées par l'environnement, telles que la prédation et la raréfaction des ressources, jouaient un rôle déterminant dans ces tendances . Ces idées ont été confortées par d'autres études, comme celle portant sur la péramorphose chez la rainette de Porto Rico . La durée de génération , ou la durée de vie de l'espèce, pourrait également être un facteur explicatif. Lorsqu'une espèce a une durée de vie relativement courte, la sélection naturelle favorise l'évolution de la pédomorphose (ex. : axolotl : 7 à 10 ans). Inversement, chez les espèces à longue durée de vie, la sélection naturelle favorise l'évolution de la péramorphose (ex. : wapiti d'Irlande : 20 à 22 ans).
Dans tout le règne animal
L'hétérochronie est responsable d'une grande variété d'effets tels que l'allongement des doigts par l'ajout de phalanges supplémentaires chez les dauphins pour former leurs nageoires, le dimorphisme sexuel , et le polymorphisme observé entre les castes d'insectes .
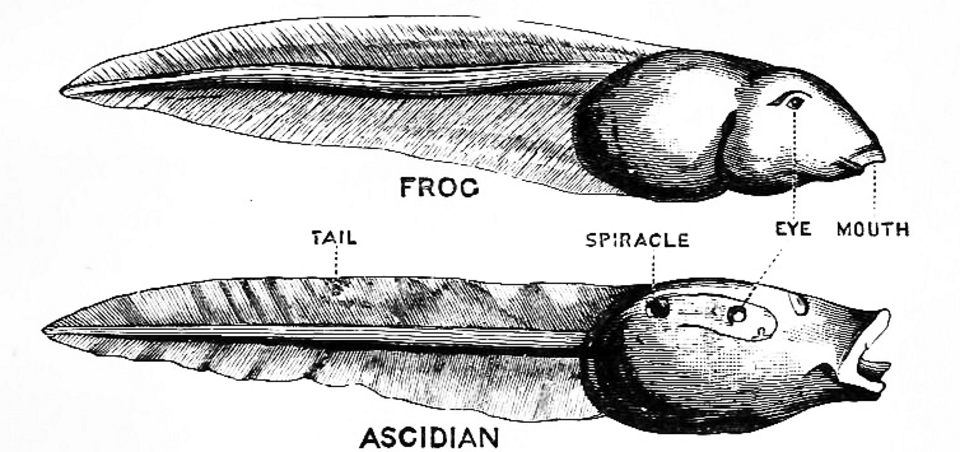
L'hypothèse de Garstang
Walter Garstang a suggéré une origine néoténique des vertébrés à partir d'une larve de tunicier , s'opposant ainsi à l'opinion de Darwin selon laquelle les tuniciers et les vertébrés descendaient d'animaux dont la forme adulte était semblable aux têtards (de grenouille) et aux larves de tuniciers. D'après Richard Dawkins , l'opinion de Garstang était également partagée par Alister Hardy , et elle l'est encore par certains biologistes modernes. Cependant, selon d'autres, des analyses génétiques plus approfondies semblent plutôt confirmer l'ancienne opinion de Darwin.
La théorie de Garstang est certes séduisante et a longtemps été largement acceptée… Malheureusement, de récentes analyses ADN ont fait pencher la balance en faveur de la théorie originale de Darwin. Si les larvacés constituent une réactualisation récente d'un scénario ancien de Garstang, ils devraient présenter une parenté plus étroite avec certaines ascidies modernes qu'avec d'autres. Hélas, il n'en est rien.
— Richard Dawkins

Chez l'homme
Concepts connexes
Le terme « hétérokairie » a été proposé en 2003 par John Spicer et Warren Burggren pour distinguer la plasticité dans le calendrier du déclenchement des événements de développement au niveau d'un individu (hétérocairie) ou d'une population (hétérochronie).