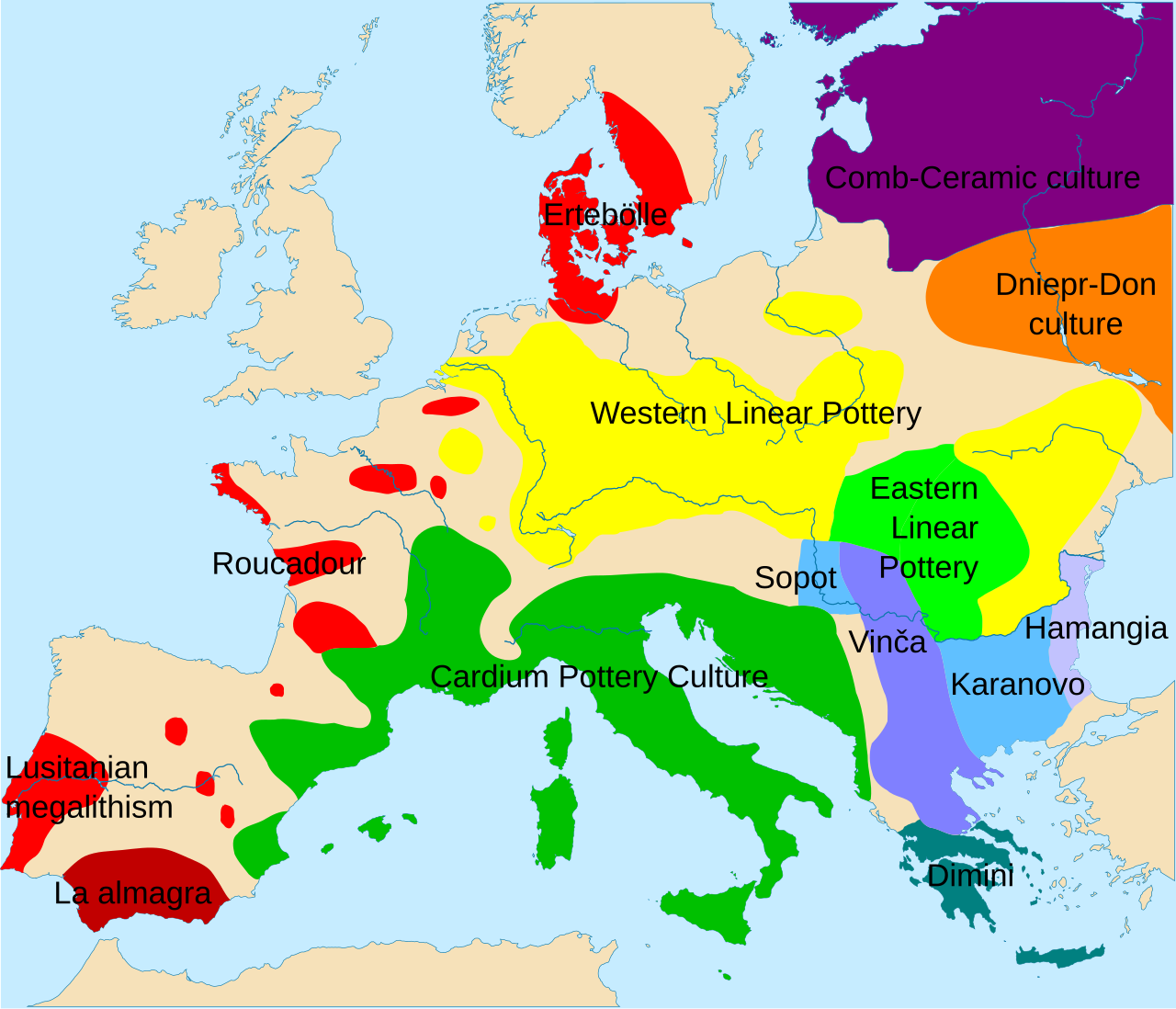
Carte montrant la fréquence de l'haplogroupe R1a en Europe L'haplogroupe R1a , ou haplogroupe R-M420 , est un haplogroupe d'ADN du chromosome Y humain qui est distribué dans une...
Worldlex WikiFrancaisLecture gratuite
Carte montrant la fréquence de l'haplogroupe R1a en Europe
Alors qu'une étude génétique indique que R1a est apparu il y a 25 000 ans, sa sous-clade M417 (R1a1a1) s'est diversifiée il y a environ 5 800 ans. Le lieu d'origine de la sous-clade joue un rôle dans le débat sur les origines des proto-indo-européens .
La mutation SNP R-M420 a été découverte après R-M17 (R1a1a), ce qui a entraîné une réorganisation de la lignée, en particulier l'établissement d'un nouveau paragroupe (désigné R-M420*) pour les lignées relativement rares qui ne sont pas dans la branche R-SRY10831.2 (R1a1) menant à R-M17.
Origines
Origines de R1a
La divergence génétique de R1a (M420) est estimée avoir eu lieu il y a 25 000 ans, soit l'époque du dernier maximum glaciaire . Une étude de 2014 menée par Peter A. Underhill et al., utilisant 16 244 individus de plus de 126 populations de toute l'Eurasie, a conclu qu'il y avait « un argument convaincant pour le Moyen-Orient, peut-être près de l'Iran actuel, comme origine géographique de hg R1a ». L'ancien enregistrement ADN a montré le premier R1a au cours du Mésolithique chez les chasseurs-cueilleurs de l'Est (d'Europe de l'Est, il y a environ 13 000 ans), et le premier cas de R* chez les anciens Eurasiens du Nord du Paléolithique supérieur , d'où les chasseurs-cueilleurs de l'Est tirent principalement leur ascendance.
Diversification de R1a1a1 (M417) et migrations anciennes
Origines de R1a (Underhill 2009 ; Origines de R1a1a (Pamjav et al. 2012) ; migration possible de R1a vers la côte baltique ; et expansion la plus ancienne et fréquence la plus élevée de R1a1a (Underhill et al. 2014)
Selon Underhill et al. (2014), la sous-clade en aval R1a-M417 s'est diversifiée en Z282 et Z93 il y a environ 5 800 ans « à proximité de l'Iran et de la Turquie orientale ». Même si R1a apparaît comme un haplogroupe du chromosome Y chez les locuteurs de diverses langues telles que le slave et l'indo-iranien , la question des origines de R1a1a est pertinente pour le débat en cours concernant l' urheimat du peuple proto-indo-européen , et peut également être pertinente pour les origines de la civilisation de la vallée de l'Indus . R1a montre une forte corrélation avec les langues indo-européennes d' Asie du Sud et de l'Ouest , d'Europe centrale et orientale et de Scandinavie étant les plus répandues en Europe de l'Est , en Asie centrale et en Asie du Sud . En Europe, Z282 est particulièrement répandu tandis qu'en Asie, Z93 domine. Le lien entre l'Y-ADN R-M17 et la propagation des langues indo-européennes a été noté pour la première fois par T. Zerjal et ses collègues en 1999.
Relation indo-européenne
Proposition de dispersion dans les steppes de R1a1a
Semino et al. (2000) ont proposé des origines ukrainiennes et une propagation postglaciaire de l'haplogroupe R1a1 pendant le maximum glaciaire tardif , amplifiée par la suite par l'expansion de la culture kourgan en Europe et vers l'est. Spencer Wells propose des origines d'Asie centrale, suggérant que la distribution et l'âge de R1a1 indiquent une migration ancienne correspondant à la propagation du peuple kourgan dans son expansion à partir de la steppe eurasienne . Selon Pamjav et al. (2012), R1a1a s'est diversifié dans les steppes eurasiennes ou dans la région du Moyen-Orient et du Caucase :
L'Asie intérieure et centrale est une zone de chevauchement pour les lignées R1a1-Z280 et R1a1-Z93 [ce qui] implique qu'une zone de différenciation précoce de R1a1-M198 s'est probablement produite quelque part dans les steppes eurasiennes ou dans la région du Moyen-Orient et du Caucase, situées entre l'Asie du Sud et l'Europe centrale et orientale.
Trois études génétiques réalisées en 2015 ont soutenu la théorie Kurgan de Gimbutas concernant l' Urheimat indo-européen . Selon ces études, les haplogroupes R1b et R1a, aujourd'hui les plus courants en Europe (R1a est également courant en Asie du Sud) se seraient développés à partir des steppes pontiques-caspiennes, en même temps que les langues indo-européennes ; elles ont également détecté une composante autosomique présente chez les Européens modernes qui n'était pas présente chez les Européens néolithiques, qui aurait été introduite avec les lignées paternelles R1b et R1a, ainsi que les langues indo-européennes.
Silva et al. (2017) ont noté que R1a en Asie du Sud « s'est probablement propagé à partir d'un seul bassin source d'Asie centrale , il semble y avoir au moins trois et probablement plus de clades fondateurs de R1a dans le sous-continent indien , ce qui correspond à de multiples vagues d'arrivée ». Selon Martin P. Richards, co-auteur de Silva et al. (2017), la prévalence de R1a en Inde était « une preuve très convaincante d'une migration substantielle de l'âge du bronze en provenance d'Asie centrale qui a très probablement amené des locuteurs indo-européens en Inde ».
Origines possibles de Yamnaya ou Corded Ware
Période du Néolithique moyen européen. Culture des peignes, vers 4200 – vers 2000 av. J.-C.Culture de la céramique cordée (vers 2900 – vers 2350 av. J.-C.)
David Anthony considère la culture Yamnaya comme l' Urheimat indo-européen . Selon Haak et al. (2015), une migration massive de la culture Yamnaya vers le nord a eu lieu vers 2 500 av. J.-C., représentant 75 % de l'ascendance génétique de la culture de la céramique cordée , notant que R1a et R1b peuvent s'être « répandus en Europe depuis l' Est après 3 000 av. J.-C. ». Pourtant, leurs sept échantillons Yamnaya appartenaient tous à la sous-clade R1b-M269 , mais aucun R1a1a n'a été trouvé dans leurs échantillons Yamnaya. Cela soulève la question de l'origine du R1a1a dans la culture de la céramique cordée, s'il ne venait pas de la culture Yamnaya.
Selon Marc Haber, l’absence de l’haplogroupe R1a-M458 en Afghanistan ne soutient pas une origine de steppe pontique-caspienne pour les lignées R1a dans les populations modernes d’Asie centrale.
Selon Leo Klejn , l'absence de l'haplogroupe R1a dans les restes de Yamnaya (malgré sa présence dans les populations énéolithiques de Samara et de chasseurs-cueilleurs de l'Est ) rend peu probable que les Européens aient hérité de l'haplogroupe R1a de Yamnaya.
L'archéologue Barry Cunliffe a déclaré que l'absence de l'haplogroupe R1a dans les spécimens Yamnaya est une faiblesse majeure dans la proposition de Haak selon laquelle R1a a une origine Yamnaya.
Semenov et Bulat (2016) plaident en faveur d'une origine Yamnaya de R1a1a dans la culture Corded Ware, notant que plusieurs publications indiquent la présence de R1a1 dans la culture Peigne Ware .
Origines sud-asiatiques proposées
Kivisild et al. (2003) ont proposé soit l' Asie du Sud soit l'Asie de l'Ouest , tandis que Mirabal et al. (2009) soutiennent à la fois l'Asie du Sud et l'Asie centrale. Sengupta et al. (2006) ont proposé des origines indiennes. Thanseem et al. (2006) ont proposé soit l'Asie du Sud soit l'Asie centrale. Sahoo et al. (2006) ont proposé soit l'Asie du Sud soit l'Asie de l'Ouest. Thangaraj et al. (2010) ont également proposé une origine sud-asiatique. Sharma et al. (2009) théorisent l'existence de R1a en Inde au-delà de 18 000 ans jusqu'à peut-être 44 000 ans.
Plusieurs études menées de 2006 à 2010 ont conclu que les populations d'Asie du Sud présentent la plus grande diversité STR au sein de R1a1a, TMRCA ultérieures plus anciennes . R1a1a est présent à la fois parmi les castes supérieures ( brahmanes ) et les castes inférieures, et bien que la fréquence soit plus élevée parmi les castes brahmanes, les datations TMRCA les plus anciennes de l'haplogroupe R1a se produisent dans la tribu Saharia , une caste répertoriée de la région de Bundelkhand en Inde centrale .
À partir de ces résultats, certains chercheurs ont avancé que R1a1a était originaire d'Asie du Sud, excluant un afflux génétique plus récent, mais mineur, de migrants indo-européens dans les régions du nord-ouest comme l'Afghanistan, le Baloutchistan, le Pendjab et le Cachemire.
La conclusion selon laquelle R1a est originaire d’Inde a été remise en question par des recherches plus récentes, apportant la preuve que R1a est arrivé en Inde avec de multiples vagues de migration.
Origines proposées de la Transcaucasie et de l'Asie occidentale et influence possible sur la civilisation de la vallée de l'Indus
Haak et al. (2015) ont découvert qu'une partie de l'ascendance Yamnaya provenait du Moyen-Orient et que les techniques néolithiques étaient probablement arrivées à la culture Yamnaya depuis les Balkans . La culture Rössen (4 600-4 300 av. J.-C.), qui était située en Allemagne et qui est antérieure à la culture de la céramique cordée, une ancienne sous-clade de R1a, à savoir L664, peut encore être trouvée.
Une partie de l'ascendance génétique sud-asiatique provient des populations d'Eurasie occidentale, et certains chercheurs ont suggéré que Z93 pourrait être arrivé en Inde via l'Iran et s'y être développé pendant la civilisation de la vallée de l'Indus .
Mascarenhas et al. (2015) ont proposé que les racines de Z93 se trouvent en Asie occidentale et ont suggéré que « Z93 et L342.2 se sont étendus dans une direction sud-est de la Transcaucasie vers l'Asie du Sud », notant qu'une telle expansion est compatible avec « les archives archéologiques de l'expansion vers l'est des populations d'Asie occidentale au 4e millénaire avant notre ère, culminant dans les migrations dites Kura-Araxes dans la période post-Uruk IV ». Pourtant, Lazaridis a noté que l'échantillon I1635 de Lazaridis et al. (2016), leur échantillon arménien Kura-Araxes, portait l'haplogroupe Y R1 b 1-M415(xM269) (également appelé R1b1a1b-CTS3187).
Selon Underhill et al. (2014), la diversification de Z93 et « l'urbanisation précoce dans la vallée de l'Indus... se sont produites il y a [5 600 ans] et la distribution géographique de R1a-M780 (Figure 3d ) peut refléter cela. » Poznik et al. (2016) notent que des « expansions marquantes » se sont produites dans R1a-Z93 il y a environ 4 500 à 4 000 ans, ce qui « précède de quelques siècles l'effondrement de la civilisation de la vallée de l'Indus. »
Cependant, selon Narasimhan et al. (2018), les éleveurs de steppes sont une source probable de R1a en Inde.
Phylogénie
L'arbre généalogique R1a comporte désormais trois niveaux principaux de ramification, avec le plus grand nombre de sous-clades définis au sein de la branche dominante et la plus connue, R1a1a (que l'on retrouvera sous divers noms tels que « R1a1 » dans la littérature relativement récente mais pas la plus récente).
Topologie
La topologie de R1a est la suivante (codes [entre parenthèses] codes non-isogg) : Tatiana et al. (2014) « le processus de diversification rapide de K-M526 s'est probablement produit en Asie du Sud-Est , avec des expansions ultérieures vers l'ouest des ancêtres des haplogroupes R et Q. »
R1a se distingue par plusieurs marqueurs uniques, notamment la mutation M420. Il s'agit d'un sous-clade de l'haplogroupe R-M173 (anciennement appelé R1). R1a a les sous-clades sœurs Haplogroupe R1b -M343 et le paragroupe R-M173*.
R-M420 (R1a)
R1a, défini par la mutation M420, possède deux branches primaires : R-M459 (R1a1) et R-YP4141 (R1a2).
En 2024, il n'existe aucun exemple connu de R1a* basal. Lorsque des exemples négatifs pour M-459 ont été découverts pour la première fois, ils ont d'abord été considérés comme un paragroupe basal rare , sous R-M420* et défini par la mutation SRY1532.2. Des exemples de R1a initialement considérés comme basaux et constituant un paragroupe sont maintenant connus pour avoir fait partie d'une bifurcation fondamentale dans R1a*, c'est-à-dire R1a2 (R-YP4141). (Le SNP SRY1532.2 précédemment défini est maintenant considéré comme peu fiable.) R1a2 a deux sous-branches : R1a2a (R-YP5018) et R1a2b (R-YP4132).
R-YP4141 (R1a2)
R1a2 (R-YP4141) possède deux branches R1a2a (R-YP5018) et R1a2b (R-YP4132).
Ce sous-clade primaire rare était initialement considéré comme faisant partie d'un paragroupe de R1a*, défini par SRY1532.2 (et censé exclure toujours M459 et ses synonymes SRY10831.2, M448, L122 et M516).
Français Le YP4141 a remplacé plus tard le SRY1532.2 – qui s'est avéré peu fiable – et le groupe R1a(xR-M459) a été redéfini comme R1a2. Il est relativement inhabituel, bien qu'il ait été testé dans plus d'une étude. Sahoo et al. (2006) ont signalé le R-SRY1532.2* pour 1/15 des échantillons Rajput de l'Himachal Pradesh . Underhill et al. (2009) ont signalé 1/51 en Norvège , 3/305 en Suède , 1/57 Macédoniens grecs , 1/150 (ou 2/150) Iraniens, 2/734 Arméniens de souche , 1/141 Kabardes , 1/121 Omanais , 1/164 aux Émirats arabes unis et 3/612 en Turquie . Les tests effectués sur 7 224 autres mâles dans 73 autres populations eurasiennes n'ont montré aucun signe de cette catégorie.
R-M459 (R1a1)
La sous-clade majeure R-M459 comprend une majorité écrasante d'individus appartenant au groupe R1a au sens large. Cependant, en 2024, tous les individus connus porteurs du M459 appartiennent au groupe R1a1a ou R1a1b ; aucun exemple de R1a1* n'a encore été identifié.
R-YP1272 (R1a1b)
R-YP1272, également connu sous le nom de R-M459(xM198), est une sous-clade primaire extrêmement rare de R1a1. Elle a été trouvée chez trois individus, originaires respectivement de Biélorussie, de Tunisie et de la communauté copte d'Égypte .
R-M17/M198 (R1a1a)
Les SNP suivants sont associés à R1a1a :
R-M417 (R1a1a1)
R1a1a1 (R-M417) est la sous-clade la plus largement trouvée, dans deux variantes que l'on trouve respectivement en Europe (R1a1a1b1 (R-Z282) ([R1a1a1a*] (R-Z282) (Underhill 2014) ) et en Asie centrale et du Sud (R1a1a1b2 (R-Z93) ([R1a1a2*] (R-Z93) Underhill 2014) ).
R-Z282 (R1a1a1b1a) (Europe de l'Est)
Cette grande sous-clade semble englober la plupart des R1a1a trouvés en Europe.
R1a1a1b1a [R1a1a1a* (Underhill (2014))] (R-Z282*) est présent dans le nord de l'Ukraine, en Biélorussie et en Russie à une fréquence d'environ 20 %.
R1a1a1b1a3 [R1a1a1a1 (Underhill (2014))] (R-Z284) est présent dans le nord-ouest de l'Europe et culmine à environ 20 % en Norvège.
R1a1a1c (M64.2, M87, M204) est apparemment rare : il a été trouvé chez 1 des 117 mâles typés dans le sud de l'Iran.
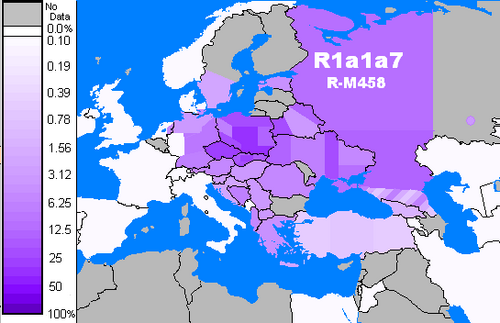
R-M458 (R1a1a1b1a1)
Distribution de fréquence du R-M458
R-M458 est un SNP principalement slave , caractérisé par sa propre mutation, et a d'abord été appelé cluster N. Underhill et al. (2009) ont découvert qu'il était présent dans les populations européennes modernes à peu près entre le bassin versant du Rhin et les montagnes de l'Oural et l'ont attribué à « un effet fondateur qui... tombe dans la période de l'Holocène précoce, 7,9 ± 2,6 KYA. » (vitesses de Zhivotovsky, 3x surévaluées) M458 a été trouvé dans un squelette d'un champ funéraire du 14e siècle à Usedom , Mecklembourg-Poméranie-Occidentale, Allemagne. L'article d'Underhill et al. (2009) rapporte également une fréquence étonnamment élevée de M458 dans certaines populations du Caucase du Nord (18 % chez les Ak Nogai , 7,8 % chez les Qara Nogai et 3,4 % chez les Abazas ).
R-L260 (R1a1a1b1a1a)
R1a1a1b1a1a (R-L260), communément appelé slave occidental ou polonais , est une sous-clade du groupe parent plus large R-M458, et a été identifié pour la première fois comme un groupe STR par Pawlowski et al. 2002. En 2010, il a été vérifié qu'il s'agissait d'un haplogroupe identifié par sa propre mutation (SNP). Il représente apparemment environ 8 % des hommes polonais, ce qui en fait le sous-clade le plus courant en Pologne. En dehors de la Pologne, il est moins courant. En plus de la Pologne, on le trouve principalement en République tchèque et en Slovaquie , et il est considéré comme « clairement slave occidental ». On estime que l'ancêtre fondateur de R-L260 a vécu entre 2000 et 3000 ans, c'est-à-dire pendant l' âge du fer , avec une expansion démographique significative il y a moins de 1 500 ans.
R-M334
R-M334 ([R1a1a1g1], un sous-clade de [R1a1a1g] (M458) cq R1a1a1b1a1 (M458) ) a été trouvé par Underhill et al. (2009) seulement chez un homme estonien et peut définir un clade très récemment fondé et de petite taille.
R1a1a1b1a2 (S466/Z280, S204/Z91)
R1a1a1b1a2b3* (Groupe K de Gwozdz)
R1a1a1b1a2b3* (M417+, Z645+, Z283+, Z282+, Z280+, CTS1211+, CTS3402, Y33+, CTS3318+, Y2613+) (cluster K de Gwozdz) est un groupe basé sur STR qui est R-M17(xM458). Ce cluster est commun en Pologne mais n'est pas exclusif à la Pologne.
R1a1a1b1a2b3a (R-L365)
R1a1a1b1a2b3a (R-L365) a été appelé au début Cluster G. [
Le R-Z93* ou R1a1a1b2* (R1a1a2* dans Underhill (2014)) est le plus courant (> 30 %) dans la région de l'Altaï sibérien du Sud en Russie, apparaissant au Kirghizistan (6 %) et dans toutes les populations iraniennes (1 à 8 %).
Le R-Z2125 est présent à des fréquences plus élevées au Kirghizistan et chez les Pachtounes afghans (> 40 %). À une fréquence de > 10 %, il est également observé dans d'autres groupes ethniques afghans et dans certaines populations du Caucase et d'Iran.
Le R-M434 est un sous-clade du Z2125. Il a été détecté chez 14 personnes (sur 3667 personnes testées), toutes dans une zone géographique restreinte allant du Pakistan à Oman . Cela reflète probablement un événement de mutation récent au Pakistan.
Le R-M560 est très rare et n'a été observé que dans quatre échantillons : deux locuteurs de Burushaski (nord du Pakistan), un locuteur de Hazara (Afghanistan) et un locuteur d'Azerbaïdjan iranien.
R-M780 est présent à une fréquence élevée en Asie du Sud : Inde, Pakistan, Afghanistan et Himalaya. La Turquie partage les sous-lignées R1a (12,1 %). Les Roms de Slovaquie partagent 3 % de R1a Le groupe est également présent à > 3 % dans certaines populations iraniennes et est présent à > 30 % chez les Roms de Croatie et de Hongrie.
Répartition géographique de R1a1a
Répartition de R1a (violet) et R1b (rouge)
Préhistorique
Français En Europe mésolithique, R1a est caractéristique des chasseurs-cueilleurs de l'Est (EHG). Un EHG mâle de la culture Veretye enterré à Peschanitsa près du lac Lacha dans l'oblast d'Arkhangelsk , en Russie vers 10 700 av. J.-C. s'est avéré porteur de l'haplogroupe paternel R1a5-YP1301 et de l'haplogroupe maternel U4a . Un mâle, nommé PES001, de Peschanitsa dans le nord-ouest de la Russie s'est avéré porteur de R1a5, et date d'il y a au moins 10 600 ans. D'autres exemples incluent les mâles Minino II (V) et Minino II (I/1), le premier portant respectivement R1a1 et le second R1a, le premier étant âgé de 10 600 ans et le second d'au moins 10 400 ans respectivement, tous deux de Minino dans le nord-ouest de la Russie. Un homme du Mésolithique de Carélie d'environ 8 800 à 7 950 av. J.-C. a été trouvé porteur de l'haplogroupe R1a. Un homme du Mésolithique enterré à Deriivka d'environ 7 000 à 6 700 av. J.-C. portait l'haplogroupe paternel R1a et l'haplogroupe maternel U5a2a . Un autre homme de Carélie d'environ 5 500 à 5 000 av. J.-C., qui était considéré comme un EHG, portait l'haplogroupe R1a. Un homme de la culture de la céramique en peigne à Kudruküla d'environ 5 900 à 3 800 av. J.-C. a été déterminé comme étant porteur de R1a et de l'haplogroupe maternel U2e1 . Selon l'archéologue David Anthony, le R1a-Z93 paternel a été trouvé à la rivière Oskol près d'un kolkhoze "Alexandrie" aujourd'hui existant, en Ukraine vers 1900. 4000 avant J.-C., « le plus ancien échantillon connu montrant l'adaptation génétique à la persistance de la lactase (13910-T) ». R1a a été trouvé dans la culture de la céramique cordée , dans laquelle il est prédominant. culture de Fatyanovo de l'âge du bronze appartiennent entièrement à R1a, en particulier au sous-clade R1a-Z93.
L'haplogroupe R1a a été retrouvé plus tard dans des fossiles anciens associés à la culture des champs d'urnes , ainsi que dans l'enterrement des restes des cultures de Sintashta , d'Andronovo , de Pazyryk , de Tagar , de Tashtyk et de Srubnaya , les habitants de l'ancienne Tanais , dans les momies du Tarim et dans l'aristocratie de Xiongnu . Les restes squelettiques d'un père et de ses deux fils, provenant d'un site archéologique découvert en 2005 près d'Eulau (en Saxe-Anhalt , en Allemagne ) et datés d'environ 2600 av. J.-C., ont été testés positifs pour le marqueur Y-SNP SRY10831.2. Le numéro Ysearch pour les restes d'Eulau est 2C46S. Le clade ancestral était donc présent en Europe il y a au moins 4 600 ans, en association avec un site de la culture très répandue de la céramique cordée .
Europe
Français En Europe, le sous-clade R1a1a est principalement caractéristique des populations balto-slaves, à deux exceptions près : les Slaves du sud et les Russes du nord. La fréquence la plus élevée de R1a1a en Europe est observée chez les Sorabes (63 %), un groupe ethnique slave occidental , suivi des Hongrois (60 %). D'autres groupes avec R1a1a important, allant de 27 % à 58 %, comprennent les Tchèques , les Polonais , les Slovènes , les Slovaques , les Moldaves , les Biélorusses , les Ruthènes , les Ukrainiens et les Russes . La fréquence de R1a diminue dans les populations du nord-est de la Russie jusqu'à 20 à 30 %, contrairement à la Russie centrale et méridionale, où sa fréquence est deux fois plus élevée. Dans les États baltes , les fréquences de R1a1a diminuent de la Lituanie (45 %) à l'Estonie (environ 30 %).
On observe également une présence significative chez les peuples d' origine germanique , avec les niveaux les plus élevés en Norvège , en Suède et en Islande , où entre 20 et 30 % des hommes sont en R1a1a. Les Vikings et les Normands ont peut-être également porté la lignée R1a1a plus loin, ce qui explique au moins une partie de la faible présence dans les îles britanniques , les îles Canaries et la Sicile . L'haplogroupe R1a1a se situe en moyenne entre 10 et 30 % chez les Allemands , avec un pic à Rostock à 31,3 %. R1a1a se trouve à une fréquence très faible chez les Néerlandais (3,7 %) et est pratiquement absent chez les Danois .
En Europe du Sud, R1a1a n'est pas courant, mais des niveaux significatifs ont été trouvés dans certaines zones, comme dans la vallée du Pas dans le nord de l'Espagne , dans certaines régions de Venise et en Calabre en Italie . Les Balkans présentent une grande variation entre les zones présentant des niveaux significatifs de R1a1a, par exemple 36 à 39 % en Slovénie , 27 à 34 % en Croatie , et plus de 30 % en Macédoine grecque , mais moins de 10 % en Albanie , au Kosovo et dans certaines parties de la Grèce au sud des gorges de l'Olympe.
R1a est pratiquement composé uniquement de la sous-clade Z284 en Scandinavie . En Slovénie, la sous-clade principale est Z282 (Z280 et M458), bien que la sous-clade Z284 ait été trouvée dans un échantillon d'un Slovène. Il y a une représentation négligeable de Z93 en Turquie , 12,1% Les Slaves occidentaux et les Hongrois sont caractérisés par une fréquence élevée de la sous-clade M458 et une faible Z92, une sous-clade de Z280. Des centaines d'échantillons slovènes et tchèques n'ont pas la sous-clade Z92 de Z280, tandis que les Polonais, les Slovaques, les Croates et les Hongrois ne présentent qu'une très faible fréquence de Z92. Les Baltes , les Slaves de l'Est , les Serbes , les Macédoniens , les Bulgares et les Roumains présentent un rapport Z280>M458 et une part élevée, voire prédominante, de Z92. Les Baltes et les Slaves de l'Est ont les mêmes sous-clades et des fréquences similaires dans une phylogénie plus détaillée des sous-clades. Le généticien russe Oleg Balanovsky a émis l'hypothèse qu'il existe une prédominance du substrat pré-slave assimilé dans la génétique des populations slaves de l'Est et de l'Ouest. Selon lui, la structure génétique commune qui distingue les Slaves de l'Est et les Baltes des autres populations peut suggérer l'explication selon laquelle le substrat pré-slave des Slaves de l'Est et de l'Ouest était principalement constitué de locuteurs baltes, qui à un moment donné ont précédé les Slaves dans les cultures de la steppe eurasienne selon les références archéologiques et toponymiques.
Haber et al. (2012) ont trouvé R1a1a-M17 dans 26,0 % (53/204) d'un ensemble d'échantillons provenant d' Afghanistan , dont 60 % (3/5) d'un échantillon de Nouristanis , 51,0 % (25/49) d'un échantillon de Pachtounes, 30,4 % (17/56) d'un échantillon de Tadjiks, 17,6 % (3/17) d'un échantillon d'Ouzbeks, 6,7 % (4/60) d'un échantillon de Hazaras, et chez le seul individu turkmène échantillonné.
Di Cristofaro et al. (2013) ont trouvé R1a1a-M198/M17 dans 56,3 % (49/87) d'une paire d'échantillons de Pachtounes d'Afghanistan (dont 20/34 ou 58,8 % d'un échantillon de Pachtounes de Baghlan et 29/53 ou 54,7 % d'un échantillon de Pachtounes de Kunduz ), 29,1 % (37/127) d'un ensemble d'échantillons d'Ouzbeks d'Afghanistan (dont 28/94 ou 29,8 % d'un échantillon d'Ouzbeks de Jawzjan , 8/28 ou 28,6 % d'un échantillon d'Ouzbeks de Sar-e Pol et 1/5 ou 20 % d'un échantillon d'Ouzbeks de Balkh ), 27,5 % (39/142) d'un ensemble d'échantillons de Tadjiks d'Afghanistan (dont 22/54 ou 40,7 % d'un échantillon de Tadjiks de Balkh , 9/35 ou 25,7 % d'un échantillon de Tadjiks de Takhar , 4/16 ou 25,0 % d'un échantillon de Tadjiks de Samangan et 4/37 ou 10,8 % d'un échantillon de Tadjiks de Badakhshan ), 16,2 % (12/74) d'un échantillon de Turkmènes de Jawzjan et 9,1 % (7/77) d'une paire d'échantillons de Hazaras d'Afghanistan (dont 7/69 ou 10,1 % d'un échantillon de Hazaras de Bamiyan et 0/8 ou 0 % d'un échantillon de Hazaras de Balkh ).
Malyarchuk et coll. (2013) ont trouvé R1a1-SRY10831.2 dans 30,0 % (12/40) d'un échantillon de Tadjiks du Tadjikistan.
Ashirbekov et al. (2017) ont trouvé R1a-M198 dans 6,03 % (78/1294) d'un ensemble d'échantillons de Kazakhs du Kazakhstan . R1a-M198 a été observé avec une fréquence supérieure à la moyenne dans les échantillons de l'étude des tribus kazakhes suivantes : 13/41 = 31,7 % d'un échantillon de Suan, 8/29 = 27,6 % d'un échantillon d'Oshaqty, 6/30 = 20,0 % d'un échantillon de Qozha, 4/29 = 13,8 % d'un échantillon de Qypshaq, 1/8 = 12,5 % d'un échantillon de Tore, 9/86 = 10,5 % d'un échantillon de Jetyru, 4/50 = 8,0 % d'un échantillon d'Argyn, 1/13 = 7,7 % d'un échantillon de Shanyshqyly, 8/122 = 6,6 % d'un échantillon d'Alimuly, 3/46 = 6,5 % d'un échantillon d'Alban. Le gène R1a-M198 a également été observé chez 5/42 = 11,9 % d'un échantillon de Kazakhs d'appartenance tribale non déclarée.
Asie du Sud
En Asie du Sud, R1a1a a souvent été observé dans un certain nombre de groupes démographiques.
En outre, les études montrent des pourcentages élevés dans des groupes régionaux diversifiés tels que les Manipuris (50 %) à l'extrême nord-est et parmi les Pendjabis (47 %) à l'extrême nord-ouest.
Un article chinois publié en 2018 a trouvé R1a-Z94 dans 38,5 % (15/39) d'un échantillon d'Ouïghours Keriyalik du village de Darya Boyi/Darya Boye, comté de Yutian , Xinjiang (于田县达里雅布依乡), R1a-Z93. dans 28,9% (22/76) d'un échantillon de Ouïghours Dolan du canton de Horiqol, comté d'Awat , Xinjiang (阿瓦提县乌鲁却勒镇), et R1a-Z93 dans 6,3 % (4/64) d'un échantillon de Ouïghours Loplik du village de Karquga/Qarchugha, comté de Yuli , Xinjiang (尉犁县喀尔曲尕乡). R1a(xZ93) n'a été observé que chez un seul des 76 Ouïghours de Dolan. Notez que le village de Darya Boyi est situé dans une oasis isolée formée par la rivière Keriya dans le désert du Taklamakan . Une étude de l'ADN-Y de 2011 a trouvé l'ADN-Y R1a1 dans 10 % d'un échantillon de Hui du sud du Yunnan, 1,6 % d'un échantillon de Tibétain du Tibet ( région autonome du Tibet ), 1,6 % d'un échantillon de Xibe du Xinjiang, 3,2 % d'un échantillon de Hui du nord du Ningxia , 9,4% d'un échantillon de Hazak ( Kazakhs ) du Xinjiang, et des taux de 24,0%, 22,2%, 35,2%, 29,2% dans 4 échantillons différents d'Ouïghours du Xinjiang, 9,1% dans un échantillon de Mongols de Mongolie intérieure . Un sous-clade différent de R1 a également été trouvé dans 1,5 % d'un échantillon de Hui du nord du Ningxia. Dans la même étude, aucun cas de R1a n'a été détecté dans 6 échantillons de Chinois Han au Yunnan , 1 échantillon de Han au Guangxi , 5 échantillons de Han au Guizhou , 2 échantillons de Han au Guangdong , 2 échantillons de Han au Fujian , 2 échantillons de Han au Zhejiang , 1 échantillon de Han à Shanghai , 1 échantillon de Han au Jiangxi , 2 échantillons de Han au Hunan , 1 échantillon de Han au Hubei , 2 échantillons de Han au Sichuan , 1 échantillon de Han à Chongqing , 3 échantillons de Han au Shandong , 5 échantillons de Han au Gansu , 3 échantillons de Han au Jilin et 2 échantillons de Han au Heilongjiang . 40 % des Salars, 45,2 % des Tadjiks du Xinjiang , 54,3 % des Dongxiang, 60,6 % des Tatars et 68,9 % des Kirghizes.au Xinjiang, dans le nord-ouest de la Chine, un échantillon testé avait R1a1-M17. Bao'an (Bonan) avait la plus grande diversité d'haplogroupes de 0,8946±0,0305 tandis que les autres minorités ethniques du nord-ouest de la Chine avaient une diversité d'haplogroupes élevée comme les Asiatiques centraux, de 0,7602±0,0546.
Les haplogroupes Y R1a-M420 et R2-M479 se retrouvent chez les peuples Ede (8,3 % et 4,2 %) et Giarai (3,7 % et 3,7 %) au Vietnam . Les Cham possèdent également les haplogroupes R-M17 (13,6 %) et R-M124 (3,4 %).
Les haplogroupes R1a1a1b2a2a (R-Z2123) et R1a1 se retrouvent respectivement chez les Khmers de Thaïlande (3,4 %) et du Cambodge (7,2 %). L'haplogroupe R1a1a1b2a1b (R-Y6) se retrouve également chez les Kuy (5 %).
Selon Changmai et al. (2022), ces fréquences d'haplogroupes proviennent des Sud-Asiatiques, qui ont laissé un héritage culturel et génétique en Asie du Sud-Est depuis le premier millénaire de notre ère.
Asie de l'Ouest
Le R1a1a a été trouvé sous diverses formes, dans la plupart des régions d' Asie occidentale , à des concentrations très variables, allant d'une présence quasi nulle dans des régions comme la Jordanie , à des niveaux beaucoup plus élevés dans certaines régions du Koweït et de l'Iran . La tribu bédouine Shimar ( Shammar ) au Koweït présente la fréquence la plus élevée au Moyen-Orient, soit 43 %.
Wells 2001 a noté que dans la partie occidentale du pays, les Iraniens présentent de faibles niveaux de R1a1a, tandis que les hommes des régions orientales de l'Iran présentent jusqu'à 35 % de R1a1a. Nasidze et al. 2004 ont trouvé R1a1a chez environ 20 % des hommes iraniens des villes de Téhéran et d'Ispahan . Regueiro 2006, dans une étude sur l'Iran , a noté des fréquences beaucoup plus élevées dans le sud que dans le nord.
Di Cristofaro et al. (2013) ont trouvé l'haplogroupe R1a dans 9,68 % (18/186) d'un ensemble d'échantillons provenant d'Iran, bien qu'avec une grande variance allant de 0 % (0/18) dans un échantillon d'Iraniens de Téhéran à 25 % (5/20) dans un échantillon d'Iraniens de Khorasan et 27 % (3/11) dans un échantillon d'Iraniens de provenance inconnue. Tous les individus iraniens R1a portaient les mutations M198 et M17, à l'exception d'un individu dans un échantillon d'Iraniens de Gilan ( n = 27), qui appartenait à R1a-SRY1532.2(xM198, M17).
Malyarchuk et al. (2013) ont trouvé R1a1-SRY10831.2 dans 20,8 % (16/77) d'un échantillon de Perses collectés dans les provinces de Khorasan et de Kerman dans l'est de l'Iran, mais ils n'ont trouvé aucun membre de cet haplogroupe dans un échantillon de 25 Kurdes collectés dans la province de Kermanshah dans l'ouest de l'Iran.
Plus au nord de ces régions d'Asie occidentale, en revanche, les niveaux de R1a1a commencent à augmenter dans le Caucase , une fois de plus de manière inégale. Plusieurs populations étudiées n'ont montré aucun signe de R1a1a, alors que les niveaux les plus élevés découverts jusqu'à présent dans la région semblent appartenir aux locuteurs de la langue karachaï-balkar parmi lesquels environ un quart des hommes testés jusqu'à présent appartiennent à l'haplogroupe R1a1a.
Dénomination historique de R1a
Le système de dénomination historique couramment utilisé pour R1a était incohérent dans différentes sources publiées, car il changeait souvent ; cela nécessite quelques explications.
En 2002, le Y Chromosome Consortium (YCC) a proposé un nouveau système de dénomination des haplogroupes (YCC 2002), qui est désormais devenu la norme. Dans ce système, les noms au format « R1 » et « R1a » sont des noms « phylogénétiques », destinés à marquer des positions dans un arbre généalogique. Les noms de mutations SNP peuvent également être utilisés pour nommer des clades ou des haplogroupes. Par exemple, comme M173 est actuellement la mutation définissant R1, R1 est également R-M173, un nom de clade « mutationnel ». Lorsqu'une nouvelle ramification dans un arbre est découverte, certains noms phylogénétiques changeront, mais par définition tous les noms mutationnels resteront les mêmes.
L'haplogroupe largement répandu défini par la mutation M17 était connu sous divers noms, tels que « Eu19 », tel qu'utilisé dans (Semino et al. 2000) dans les anciens systèmes de dénomination. La proposition YCC de 2002 a attribué le nom R1a à l'haplogroupe défini par la mutation SRY1532.2. Cela incluait Eu19 (c'est-à-dire R-M17) comme sous-clade, donc Eu19 a été nommé R1a1. Remarque, SRY1532.2 est également connu sous le nom de SRY10831.2 La découverte de M420 en 2009 a provoqué une réattribution de ces noms phylogénétiques. (Underhill et al. 2009 et ISOGG 2012) R1a est désormais défini par la mutation M420 : dans cet arbre mis à jour, le sous-clade défini par SRY1532.2 est passé de R1a à R1a1, et Eu19 (R-M17) de R1a1 à R1a1a.
Les mises à jour les plus récentes enregistrées sur la page Web de référence ISOGG concernent les branches de R-M17, y compris une branche majeure, R-M417.
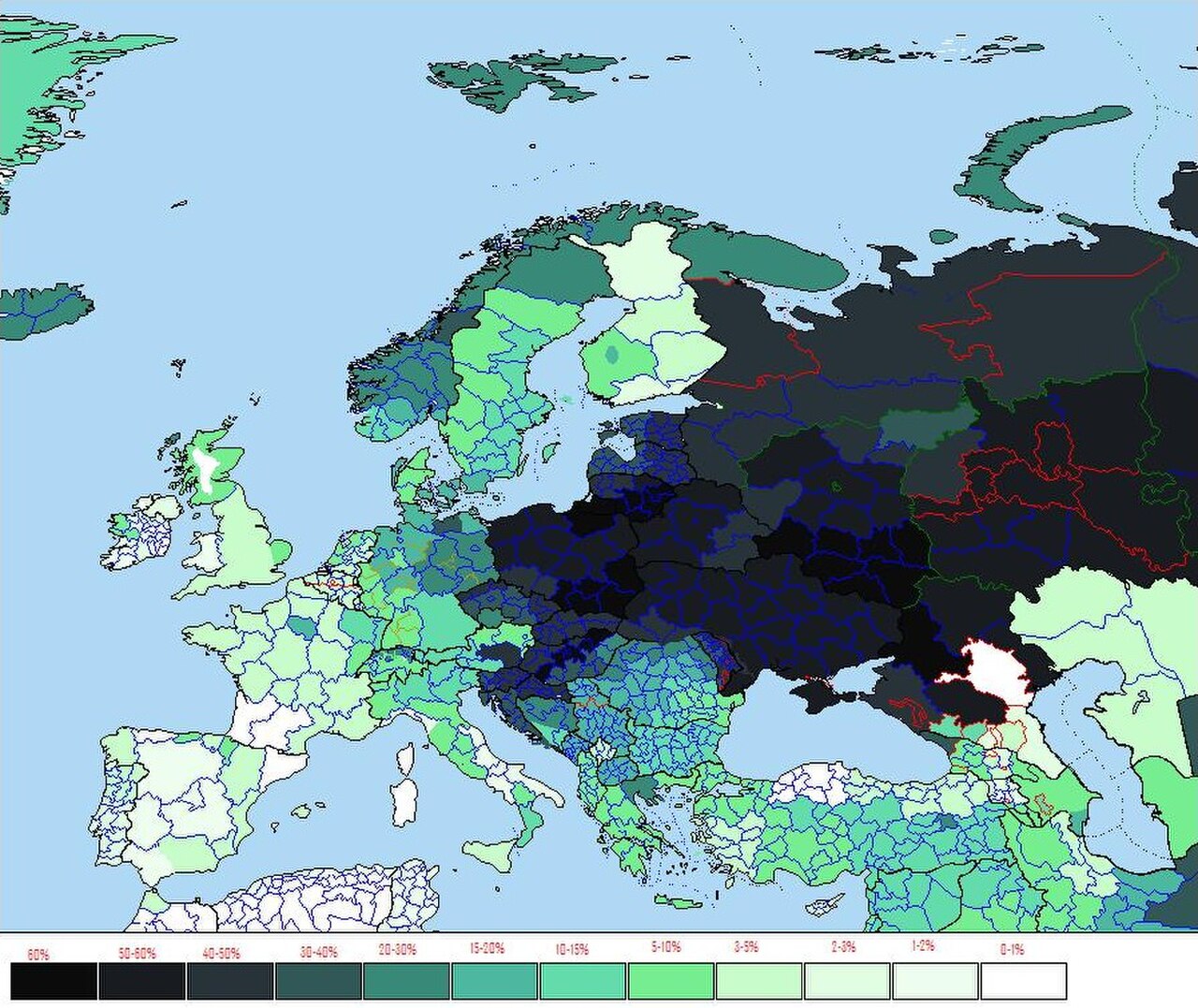
_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg/1280px-R1a_origins_(Underhill_2010)_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg)