Les séquences répétées (également appelées éléments répétitifs , unités répétitives ou répétitions ) sont des motifs courts ou longs qui apparaissent en plusieurs copies dans tout le génome . Dans de nombreux organismes, une fraction importante de l' ADN génomique est répétitive, plus des deux tiers de la séquence étant constituée d'éléments répétitifs chez l'homme. Certaines de ces séquences répétées sont nécessaires au maintien d'importantes structures du génome telles que les télomères ou les centromères .
Les séquences répétées sont classées en différentes classes en fonction de caractéristiques telles que la structure, la longueur, l'emplacement, l'origine et le mode de multiplication. La disposition des éléments répétitifs dans le génome peut consister soit en des réseaux directement adjacents appelés répétitions en tandem , soit en des répétitions dispersées dans tout le génome appelées répétitions intercalées . Les répétitions en tandem et les répétitions intercalées sont en outre classées en sous-classes en fonction de la longueur de la séquence répétée et/ou du mode de multiplication.
Si certaines séquences d'ADN répétées sont importantes pour le fonctionnement cellulaire et la maintenance du génome, d'autres séquences répétitives peuvent être nocives. De nombreuses séquences d'ADN répétitives ont été associées à des maladies humaines telles que la maladie de Huntington et l'ataxie de Friedreich. Certains éléments répétitifs sont neutres et se produisent lorsqu'il n'y a pas de sélection pour des séquences spécifiques en fonction de la façon dont se produit la transposition ou le croisement . Cependant, une abondance de répétitions neutres peut encore influencer l'évolution du génome à mesure qu'elles s'accumulent au fil du temps. Dans l'ensemble, les séquences répétées constituent un domaine d'intérêt important car elles peuvent fournir des informations sur les maladies humaines et l'évolution du génome.
Histoire
Dans les années 1950, Barbara McClintock a observé pour la première fois la transposition de l'ADN et a illustré les fonctions du centromère et du télomère au symposium de Cold Spring Harbor. Les travaux de McClintock ont ouvert la voie à la découverte des séquences répétées, car la transposition, la structure du centromère et la structure du télomère sont toutes possibles grâce à des éléments répétitifs, mais cela n'était pas entièrement compris à l'époque. Le terme « séquence répétée » a été utilisé pour la première fois par Roy John Britten et DE Kohne en 1968 ; ils ont découvert que plus de la moitié des génomes eucaryotes étaient de l'ADN répétitif grâce à leurs expériences sur la réassociation de l'ADN. Bien que les séquences d'ADN répétitives soient conservées et omniprésentes, leur rôle biologique était encore inconnu. Dans les années 1990, davantage de recherches ont été menées pour élucider la dynamique évolutive des répétitions minisatellites et microsatellites en raison de leur importance dans la criminalistique basée sur l'ADN et l'écologie moléculaire . Les répétitions dispersées dans l'ADN ont été de plus en plus reconnues comme une source potentielle de variation et de régulation génétiques . La découverte de maladies délétères liées à l'ADN répétitif a stimulé un intérêt accru pour ce domaine d'étude. Dans les années 2000, les données du séquençage complet du génome eucaryote ont permis l'identification de différents promoteurs, activateurs et ARN régulateurs qui sont tous codés par des régions répétitives. Aujourd'hui, les rôles structurels et régulateurs des séquences d'ADN répétitives restent un domaine de recherche actif.
Types et fonctions
De nombreuses séquences répétées sont probablement des restes non fonctionnels et en décomposition d' éléments transposables , ceux-ci ont été qualifiés d'ADN « indésirable » ou « égoïste ». Néanmoins, certaines répétitions peuvent parfois être exaptées pour d'autres fonctions.
Répétitions en tandem
Les répétitions en tandem sont des séquences répétées qui sont directement adjacentes les unes aux autres dans le génome. Les répétitions en tandem peuvent varier en termes de nombre de nucléotides constituant la séquence répétée, ainsi que du nombre de fois que la séquence se répète. Lorsque la séquence répétée ne mesure que 2 à 10 nucléotides de long, la répétition est appelée répétition courte en tandem (STR) ou microsatellite . Lorsque la séquence répétée mesure 10 à 60 nucléotides de long, la répétition est appelée minisatellite . [ Pour les minisatellites et les microsatellites, le nombre de fois que la séquence se répète à un seul locus peut varier de deux à des centaines de fois.
Les répétitions en tandem ont une grande variété de fonctions biologiques dans le génome. Par exemple, les minisatellites sont souvent des points chauds de recombinaison homologue méiotique dans les organismes eucaryotes. La recombinaison se produit lorsque deux chromosomes homologues s'alignent, se brisent et se rejoignent pour échanger des morceaux. La recombinaison est importante en tant que source de diversité génétique, en tant que mécanisme de réparation de l'ADN endommagé et en tant qu'étape nécessaire à la ségrégation appropriée des chromosomes lors de la méiose. La présence d'ADN à séquences répétées facilite l'alignement des zones d'homologie, contrôlant ainsi le moment et le lieu de la recombinaison.
En plus de jouer un rôle important dans la recombinaison, les répétitions en tandem jouent également des rôles structurels importants dans le génome. Par exemple, les télomères sont principalement composés de répétitions en tandem TTAGGG. quadruplexes G hautement organisées qui protègent les extrémités de l'ADN chromosomique de la dégradation. Les éléments répétitifs sont également enrichis au milieu des chromosomes. Les centromères sont les régions très compactes des chromosomes qui relient les chromatides sœurs entre elles et permettent également au fuseau mitotique de se fixer et de séparer les chromatides sœurs pendant la division cellulaire. Les centromères sont composés d'une répétition en tandem de 177 paires de bases appelée répétition α-satellite. L'hétérochromatine péricentromérique, l'ADN qui entoure le centromère et est important pour le maintien structurel, est composée d'un mélange de différentes sous-familles satellites, notamment les satellites α, β et γ ainsi que les répétitions HSATII, HSATIII et sn5.

Certaines séquences répétitives, telles que celles ayant des rôles structurels évoqués ci-dessus, jouent des rôles nécessaires au bon fonctionnement biologique. D'autres répétitions en tandem ont des rôles délétères qui entraînent des maladies. De nombreuses autres répétitions en tandem ont cependant des fonctions inconnues ou mal comprises.
Répétitions intercalées
Les répétitions intercalées sont des séquences d'ADN identiques ou similaires qui se trouvent à différents endroits du génome. Les répétitions intercalées se distinguent des répétitions en tandem en ce que les séquences répétées ne sont pas directement adjacentes les unes aux autres mais peuvent être dispersées entre différents chromosomes ou éloignées les unes des autres sur le même chromosome. La plupart des répétitions intercalées sont des éléments transposables (ET), des séquences mobiles qui peuvent être « coupées et collées » ou « copiées et collées » à différents endroits du génome. Les ET étaient à l'origine appelés « gènes sauteurs » pour leur capacité à se déplacer, mais ce terme est quelque peu trompeur car tous les ET ne sont pas des gènes discrets.
Les éléments transposables qui sont transcrits en ARN, rétrotranscrits en ADN, puis réintégrés dans le génome sont appelés rétrotransposons . Tout comme les répétitions en tandem sont subdivisées en sous-catégories en fonction de la longueur de la séquence répétitive, il existe de nombreux types différents de rétrotransposons. Les éléments nucléaires intercalés longs ( LINE ) ont généralement une longueur de 3 à 7 kilobases. Les éléments nucléaires intercalés courts ( SINE ) ont généralement une longueur de 100 à 300 paires de bases et ne dépassent pas 600 paires de bases. Les rétrotransposons à répétition terminale longue (LTR) sont une troisième grande classe de rétrotransposons et sont caractérisés par des séquences hautement répétitives aux extrémités de la répétition. Lorsqu'un élément transposable ne passe pas par l'ARN en tant qu'intermédiaire, il est appelé transposon d'ADN . D'autres systèmes de classification désignent les rétrotransposons comme des éléments transposables de « classe I » et les transposons d'ADN comme des éléments transposables de « classe II ».
Les éléments transposables sont estimés à 45 % du génome humain. Étant donné que la propagation incontrôlée des ET pourrait faire des ravages sur le génome, de nombreux mécanismes de régulation ont évolué pour faire taire leur propagation, notamment la méthylation de l'ADN, les modifications des histones, les ARN non codants (ARNnc) y compris les petits ARN interférents (ARNsi), les remodeleurs de la chromatine, les variantes d'histones et d'autres facteurs épigénétiques. Cependant, les ET jouent un grand nombre de fonctions biologiques importantes. Lorsque les ET sont introduits dans un nouvel hôte, par exemple à partir d'un virus, ils augmentent la diversité génétique. Dans certains cas, les organismes hôtes trouvent de nouvelles fonctions pour les protéines qui naissent de l'expression des ET dans un processus évolutif appelé exaptation des ET. Des recherches récentes suggèrent également que les ET servent à maintenir la structure de la chromatine d'ordre supérieur et l'organisation génomique en 3D. De plus, les ET contribuent à réguler l'expression d'autres gènes en servant d' amplificateurs distaux et de sites de liaison aux facteurs de transcription.
La prévalence des éléments intercalés dans le génome a suscité l'intérêt de recherches plus poussées sur leurs origines et leurs fonctions. Certains éléments intercalés spécifiques ont été caractérisés, tels que la répétition Alu et LINE1.
Recombinaison intrachromosomique
La recombinaison homologue entre les séquences répétées chromosomiques dans les cellules somatiques de Nicotiana tabacum s'est avérée augmentée par l'exposition à la mitomycine C , un agent alkylant bifonctionnel qui réticule les brins d'ADN. Cette augmentation de la recombinaison a été attribuée à une réparation recombinatoire intrachromosomique accrue. Par ce processus, l'ADN endommagé par la mitomycine C dans une séquence est réparé en utilisant les informations intactes de l'autre séquence répétée.
Répétitions directes et inversées
Alors que les répétitions en tandem et entrecoupées sont distinguées en fonction de leur emplacement dans le génome, les répétitions directes et inversées sont distinguées en fonction de l'ordre des bases nucléotidiques. Les répétitions directes se produisent lorsqu'une séquence nucléotidique est répétée avec la même directionnalité. Les répétitions inversées se produisent lorsqu'une séquence nucléotidique est répétée dans la direction inverse. Par exemple, une répétition directe de « CATCAT » serait une autre répétition de « CATCAT ». En revanche, la répétition inversée serait « ATGATG ». Lorsqu'il n'y a pas de nucléotides séparant la répétition inversée, comme « CATCATATGATG », la séquence est appelée répétition palindromique. Les répétitions inversées peuvent jouer des rôles structurels dans l'ADN et l'ARN en formant des boucles de tige et des cruciformes.
Séquences répétées dans les maladies humaines
Chez l'homme, certaines séquences répétées d'ADN sont associées à des maladies. Plus précisément, les séquences répétées en tandem sont à l'origine de plusieurs maladies humaines , en particulier les maladies à répétition de trinucléotides telles que la maladie de Huntington , le syndrome de l'X fragile , plusieurs ataxies spinocérébelleuses , la dystrophie myotonique et l'ataxie de Friedreich . Les expansions de répétitions de trinucléotides dans la lignée germinale au cours de générations successives peuvent conduire à des manifestations de plus en plus graves de la maladie. Ces expansions de répétitions de trinucléotides peuvent se produire par glissement de brin pendant la réplication de l'ADN ou pendant la synthèse de réparation de l'ADN . Il a été noté que les gènes contenant des répétitions CAG pathogènes codent souvent des protéines qui ont elles-mêmes un rôle dans la réponse aux dommages de l'ADN et que les expansions de répétitions peuvent altérer des voies spécifiques de réparation de l'ADN. Une réparation défectueuse des dommages de l'ADN dans les séquences répétées peut provoquer une expansion supplémentaire de ces séquences, créant ainsi un cercle vicieux de pathologie.
Maladie de Huntington
.jpg/1280px-Huntington's_disease_(5880985560).jpg)
La maladie de Huntington est une maladie neurodégénérative due à l'expansion de la séquence de trinucléotides répétés CAG dans l'exon 1 du gène de la huntingtine ( HTT ). Ce gène est responsable du codage de la protéine huntingtine qui joue un rôle dans la prévention de l'apoptose, autrement appelée mort cellulaire, et dans la réparation des dommages oxydatifs de l'ADN . Dans la maladie de Huntington, l'expansion de la séquence de trinucléotides CAG code pour une protéine huntingtine mutante avec un domaine polyglutamine étendu. Ce domaine provoque la formation d'agrégats de protéines dans les cellules nerveuses empêchant le fonctionnement cellulaire normal et entraînant une neurodégénérescence.
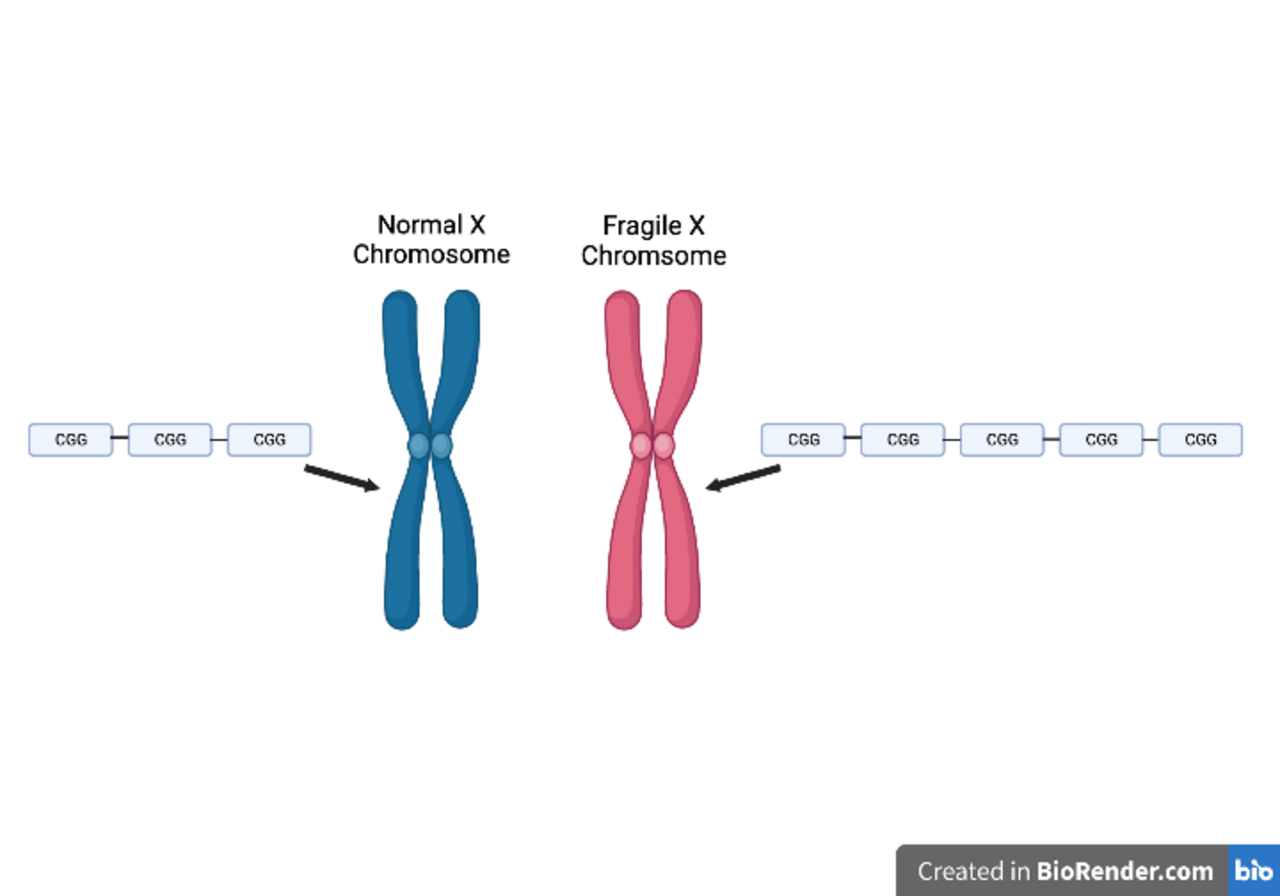
Syndrome de l'X fragile
Le syndrome de l'X fragile est causé par l'expansion de la séquence d'ADN CCG dans le gène FMR1 sur le chromosome X. Ce gène produit la protéine de liaison à l'ARN FMRP. Dans le cas du syndrome de l'X fragile, la séquence répétée rend le gène instable et donc rend silencieux le gène FMR1. Étant donné que le gène réside sur le chromosome X, les femmes qui ont deux chromosomes X sont moins affectées que les hommes qui n'ont qu'un chromosome X et un chromosome Y, car le deuxième chromosome X peut compenser le silence du gène sur l'autre chromosome X.
Ataxies spinocérébelleuses
La maladie des ataxies spinocérébelleuses présente des séquences répétées de trinucléotides CAG qui sous-tendent plusieurs types d'ataxies spinocérébelleuses (SCAs- SCA1 ; SCA2 ; SCA3 ; SCA6 ; SCA7 ; SCA12 ; SCA17 ). Comme dans la maladie de Huntington, la queue de polyglutamine créée en raison de cette expansion de trinucléotides provoque l'agrégation de protéines, empêchant la fonction cellulaire normale et provoquant une neurodégénérescence.
Ataxie de Friedreich
L'ataxie de Friedreich est un type d'ataxie qui présente une séquence de répétition étendue GAA dans le gène de la frataxine. Le gène de la frataxine est responsable de la production de la protéine frataxine, qui est une protéine mitochondriale impliquée dans la production d'énergie et la respiration cellulaire. La séquence GAA étendue entraîne le silençage du premier intron, ce qui entraîne une perte de fonction dans la protéine frataxine. La perte d'un gène FXN fonctionnel entraîne des problèmes de fonctionnement mitochondrial dans son ensemble et peut se présenter phénotypiquement chez les patients sous forme de difficultés à marcher.
Dystrophie myotonique
La dystrophie myotonique est une maladie qui se manifeste par une faiblesse musculaire et se présente sous deux formes principales : la DM1 et la DM2. Les deux types de dystrophie myotonique sont dus à des séquences d'ADN étendues. Dans la DM1, la séquence d'ADN étendue est CTG tandis que dans la DM2, c'est CCTG. Ces deux séquences se trouvent sur des gènes différents, la séquence étendue dans la DM2 se trouvant sur le gène ZNF9 et la séquence étendue dans la DM1 se trouvant sur le gène DMPK . Les deux gènes ne codent pas pour des protéines contrairement à d'autres maladies comme la maladie de Huntington ou le syndrome de l'X fragile. Il a cependant été démontré qu'il existe un lien entre la toxicité de l'ARN et les séquences répétées dans la DM1 et la DM2.
Sclérose latérale amyotrophique et démence frontotemporale
Toutes les maladies causées par des séquences répétées d'ADN ne sont pas des maladies à répétition de trinucléotides. Les maladies telles que la sclérose latérale amyotrophique et la démence frontotemporale sont causées par des séquences répétées d'hexanucléotides GGGGCC dans le gène C9orf72 , provoquant une toxicité de l'ARN qui conduit à une neurodégénérescence.
Biotechnologie
L'ADN répétitif est difficile à séquencer à l'aide des techniques de séquençage de nouvelle génération, car l'assemblage de séquences à partir de courtes lectures ne permet tout simplement pas de déterminer la longueur d'une partie répétitive. Ce problème est particulièrement grave pour les microsatellites, qui sont constitués de minuscules unités de répétition de 1 à 6 pb. Bien qu'elles soient difficiles à séquencer, ces courtes répétitions ont une grande valeur dans les empreintes génétiques et les études évolutives. De nombreux chercheurs ont historiquement laissé de côté les séquences répétitives lors de l'analyse et de la publication de données sur le génome entier en raison de limitations techniques.
Bustos et al. ont proposé une méthode de séquençage de longues séquences d'ADN répétitives. La méthode combine l'utilisation d'un vecteur linéaire pour la stabilisation et l'exonucléase III pour la suppression des régions riches en répétitions de séquences simples continues (SSR). Tout d'abord, les fragments riches en SSR sont clonés dans un vecteur linéaire qui peut incorporer de manière stable des répétitions en tandem jusqu'à 30 kb. L'expression des répétitions est interdite par les terminateurs transcriptionnels du vecteur. La deuxième étape implique l'utilisation de l'exonucléase III. L'enzyme peut supprimer le nucléotide à l'extrémité 3', ce qui entraîne la production d'une suppression unidirectionnelle des fragments SSR. Enfin, ce produit qui a supprimé des fragments est multiplié et analysé par PCR sur colonies. La séquence est ensuite construite par un séquençage ordonné d'un ensemble de clones contenant différentes suppressions.