/K+
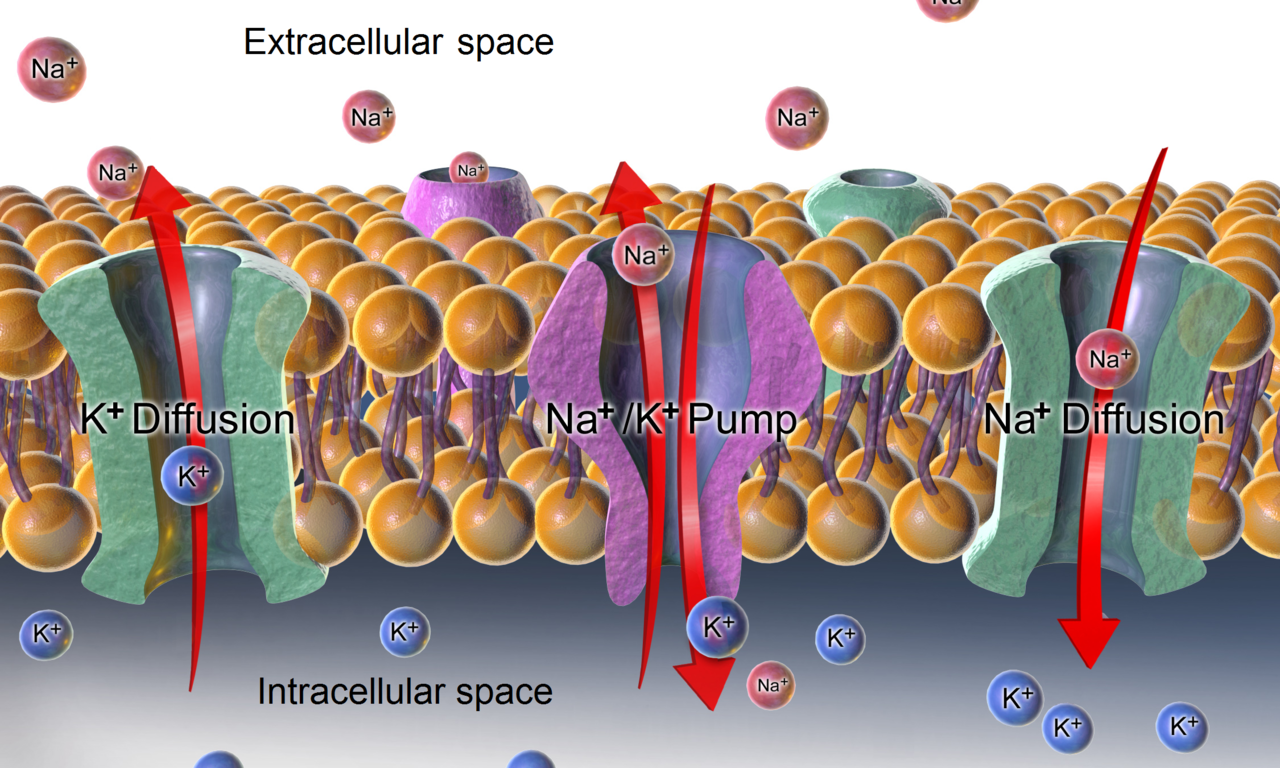
-L'ATPase , ainsi que les effets de diffusion des ions impliqués, sont des mécanismes majeurs pour maintenir le potentiel de repos à travers les membranes des cellules animales.
Le potentiel de membrane relativement statique des cellules quiescentes est appelé potentiel de membrane de repos (ou tension de repos), par opposition aux phénomènes électrochimiques dynamiques spécifiques appelés potentiel d'action et potentiel de membrane gradué . Le potentiel de membrane de repos a une valeur d'environ -70 mV ou -0,07 V.
Outre les deux derniers, qui se produisent dans les cellules excitables ( neurones , muscles et certaines cellules sécrétoires des glandes ), la tension membranaire de la majorité des cellules non excitables peut également subir des changements en réponse à des stimuli environnementaux ou intracellulaires. Le potentiel de repos existe en raison des différences de perméabilité membranaire pour les ions potassium , sodium , calcium et chlorure , qui résultent à leur tour de l'activité fonctionnelle de divers canaux ioniques , transporteurs et échangeurs d'ions. De manière conventionnelle, le potentiel de repos de la membrane peut être défini comme une valeur de base relativement stable de la tension transmembranaire dans les cellules animales et végétales.
Comme la perméabilité membranaire du potassium est bien supérieure à celle des autres ions et en raison du fort gradient chimique du potassium, les ions potassium circulent du cytosol vers l'espace extracellulaire en transportant une charge positive, jusqu'à ce que leur mouvement soit équilibré par l'accumulation d'une charge négative sur la surface interne de la membrane. Là encore, en raison de la perméabilité relative élevée du potassium, le potentiel membranaire résultant est presque toujours proche du potentiel d'inversion du potassium . Mais pour que ce processus se produise, un gradient de concentration des ions potassium doit d'abord être établi. Ce travail est effectué par les pompes/transporteurs et/ou échangeurs d'ions et est généralement alimenté par l'ATP .
Dans le cas du potentiel de membrane au repos à travers la membrane plasmique d'une cellule animale , les gradients de potassium (et de sodium) sont établis par la Na + /K + -ATPase (pompe sodium-potassium) qui transporte 2 ions potassium à l'intérieur et 3 ions sodium à l'extérieur au prix d'une molécule d'ATP. Dans d'autres cas, par exemple, un potentiel de membrane peut être établi par acidification de l'intérieur d'un compartiment membranaire (comme la pompe à protons qui génère un potentiel de membrane à travers les membranes des vésicules synaptiques ).
Électroneutralité
Dans la plupart des traitements quantitatifs du potentiel de membrane, comme la dérivation de l'équation de Goldman , on suppose l'électroneutralité , c'est-à-dire qu'il n'y a pas d'excès de charge mesurable de chaque côté de la membrane. Ainsi, bien qu'il existe un potentiel électrique à travers la membrane en raison de la séparation des charges, il n'y a pas de différence mesurable réelle dans la concentration globale d'ions positifs et négatifs à travers la membrane (comme cela est estimé ci-dessous), c'est-à-dire qu'il n'y a pas d'excès de charge mesurable réel de chaque côté. Cela se produit parce que l'effet de la charge sur le potentiel électrochimique est considérablement plus important que l'effet de la concentration, de sorte qu'un changement indétectable de concentration crée un grand changement de potentiel électrique.
Génération du potentiel de repos
Les membranes cellulaires ne sont généralement perméables qu'à un sous-ensemble d'ions. Il s'agit généralement d'ions potassium, d'ions chlorure, d'ions bicarbonate et d'autres. Pour simplifier la description de la base ionique du potentiel de repos de la membrane, il est plus utile de considérer d'abord une seule espèce ionique, puis les autres. Étant donné que les potentiels transplasmiques de la membrane sont presque toujours déterminés principalement par la perméabilité au potassium, c'est par là qu'il faut commencer.
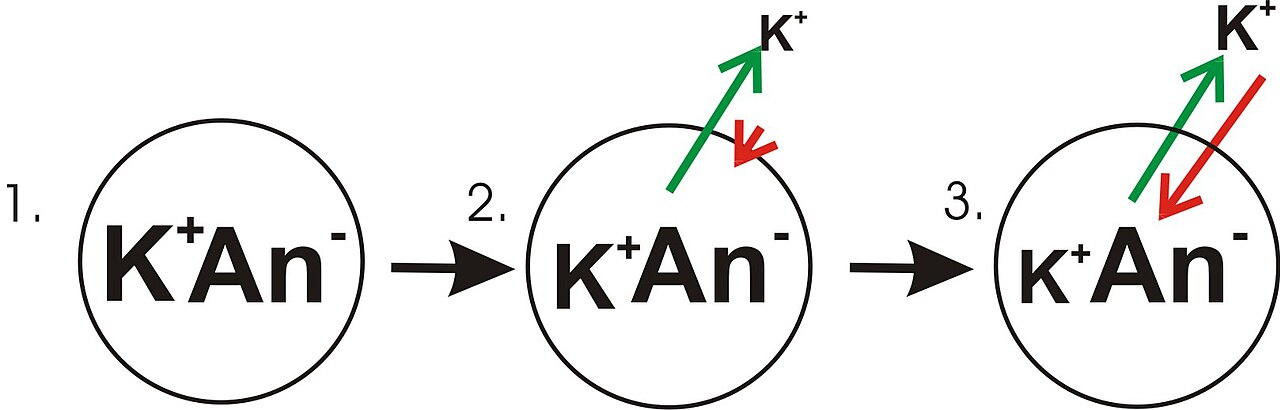
- Le panneau 1 du diagramme montre une représentation schématique d'une cellule simple où un gradient de concentration a déjà été établi. Ce panneau est dessiné comme si la membrane n'avait aucune perméabilité à aucun ion. Il n'y a pas de potentiel de membrane car malgré l'existence d'un gradient de concentration pour le potassium, il n'y a pas de déséquilibre de charge net à travers la membrane. Si la membrane devenait perméable à un type d'ion plus concentré d'un côté de la membrane, cet ion contribuerait alors à la tension de la membrane car les ions perméants se déplaceraient à travers la membrane avec un mouvement net de ce type d'ion vers le bas du gradient de concentration. Il y aurait un mouvement net du côté de la membrane avec une concentration plus élevée de l'ion vers le côté avec une concentration plus faible. Un tel mouvement d'un ion à travers la membrane entraînerait un déséquilibre net de charge à travers la membrane et un potentiel de membrane. Il s'agit d'un mécanisme courant par lequel de nombreuses cellules établissent un potentiel de membrane.
- Dans le panneau 2 du diagramme, la membrane cellulaire a été rendue perméable aux ions potassium, mais pas aux anions (An − ) à l'intérieur de la cellule. Ces anions sont principalement apportés par les protéines. Il y a de l'énergie stockée dans le gradient de concentration en ions potassium qui peut être convertie en gradient électrique lorsque les ions potassium (K + ) sortent de la cellule. Notez que les ions potassium peuvent traverser la membrane dans les deux sens, mais par le processus purement statistique qui découle de la concentration plus élevée d'ions potassium à l'intérieur de la cellule, il y aura plus d'ions potassium sortant de la cellule. Comme il y a une concentration plus élevée d'ions potassium à l'intérieur des cellules, leur mouvement moléculaire aléatoire est plus susceptible de rencontrer le pore de perméabilité ( canal ionique ), ce qui est le cas pour les ions potassium qui sont à l'extérieur et à une concentration plus faible. Un K + interne est simplement « plus susceptible » de quitter la cellule qu'un K + extracellulaire d'y entrer. Il s'agit d'une question de diffusion effectuant un travail en dissipant le gradient de concentration. Lorsque le potassium quitte la cellule, il laisse derrière lui les anions. Par conséquent, une séparation de charge se développe lorsque K + quitte la cellule. Cette séparation de charge crée une tension transmembranaire. Cette tension transmembranaire est le potentiel de membrane. Au fur et à mesure que le potassium continue de quitter la cellule, séparant davantage de charges, le potentiel de membrane continue de croître. La longueur des flèches (verte indiquant le gradient de concentration, rouge indiquant la tension) représente l'ampleur du mouvement des ions potassium dû à chaque forme d'énergie. La direction de la flèche indique la direction dans laquelle cette force particulière est appliquée. Ainsi, la tension membranaire qui se forme est une force croissante qui agit à l'encontre de la tendance au mouvement net des ions potassium vers le bas du gradient de concentration en potassium.
- Dans le panneau 3, la tension membranaire a augmenté au point que sa « force » correspond désormais aux gradients de concentration. Étant donné que ces forces (qui sont appliquées à K + ) ont maintenant la même force et sont orientées dans des directions opposées, le système est maintenant en équilibre . En d'autres termes, la tendance du potassium à quitter la cellule en descendant son gradient de concentration correspond maintenant à la tendance de la tension membranaire à ramener les ions potassium dans la cellule. K + continue de se déplacer à travers la membrane, mais la vitesse à laquelle il entre et sort de la cellule est la même, il n'y a donc pas de courant potassique net . Comme K + est à l'équilibre, le potentiel membranaire est stable, ou « au repos » (EK K ).
La tension de repos est le résultat de plusieurs enzymes de translocation d'ions ( uniporteurs , cotransporteurs et pompes ) dans la membrane plasmique, fonctionnant en parallèle de manière constante, chaque translocateur d'ions ayant sa force électromotrice caractéristique (= potentiel d'inversion = 'tension d'équilibre'), en fonction des concentrations particulières de substrat à l'intérieur et à l'extérieur ( ATP interne inclus dans le cas de certaines pompes). L'ATPase exportatrice d'H + rend la tension membranaire dans les plantes et les champignons beaucoup plus négative que dans les cellules animales plus étudiées, où la tension de repos est principalement déterminée par des canaux ioniques sélectifs.
Dans la plupart des neurones, le potentiel de repos a une valeur d'environ −70 mV. Le potentiel de repos est principalement déterminé par les concentrations d' ions dans les fluides des deux côtés de la membrane cellulaire et par les protéines de transport d'ions présentes dans la membrane cellulaire. La manière dont les concentrations d'ions et les protéines de transport membranaire influencent la valeur du potentiel de repos est décrite ci-dessous.
Le potentiel de repos d'une cellule peut être mieux compris en le considérant en termes de potentiels d'équilibre. Dans le diagramme d'exemple ici, la cellule modèle n'a reçu qu'un seul ion perméant (le potassium). Dans ce cas, le potentiel de repos de cette cellule serait le même que le potentiel d'équilibre du potassium.
Cependant, une cellule réelle est plus compliquée, ayant des perméabilités à de nombreux ions, chacun d'eux contribuant au potentiel de repos. Pour mieux comprendre, considérons une cellule avec seulement deux ions perméants, le potassium et le sodium. Considérons un cas où ces deux ions ont des gradients de concentration égaux dirigés dans des directions opposées, et où les perméabilités membranaires aux deux ions sont égales. K + quittant la cellule aura tendance à entraîner le potentiel membranaire vers E K . Na + entrant dans la cellule aura tendance à entraîner le potentiel membranaire vers le potentiel d'inversion du sodium E Na . Étant donné que les perméabilités aux deux ions ont été définies comme égales, le potentiel membranaire finira, à la fin du bras de fer Na + /K + , à mi-chemin entre E Na et E K . Comme E Na et E K étaient égaux mais de signes opposés, le milieu entre les deux est nul, ce qui signifie que la membrane reposera à 0 mV.
Notez que même si le potentiel de membrane à 0 mV est stable, il ne s'agit pas d'une condition d'équilibre car aucun des ions contributeurs n'est en équilibre. Les ions diffusent le long de leurs gradients électrochimiques à travers les canaux ioniques, mais le potentiel de membrane est maintenu par un afflux continu de K + et un afflux de Na + via des transporteurs d'ions . Une telle situation avec des perméabilités similaires pour les ions antagonistes, comme le potassium et le sodium dans les cellules animales, peut être extrêmement coûteuse pour la cellule si ces perméabilités sont relativement importantes, car il faut beaucoup d' énergie ATP pour pomper les ions. Étant donné qu'aucune cellule réelle ne peut se permettre des perméabilités ioniques aussi égales et aussi importantes au repos, le potentiel de repos des cellules animales est déterminé par une perméabilité élevée prédominante au potassium et ajusté à la valeur requise en modulant les perméabilités et les gradients de sodium et de chlorure.
Dans une cellule animale saine, la perméabilité au Na + est d'environ 5 % de la perméabilité au K + , voire moins, alors que les potentiels d'inversion respectifs sont de +60 mV pour le sodium ( E Na ) et de −80 mV pour le potassium ( E K ). Ainsi, le potentiel de membrane ne sera pas exactement à E K , mais plutôt dépolarisé par rapport à E K d'une quantité d'environ 5 % de la différence de 140 mV entre E K et E Na . Ainsi, le potentiel de repos de la cellule sera d'environ −73 mV.
Dans une notation plus formelle, le potentiel de membrane est la moyenne pondérée du potentiel d'équilibre de chaque ion contribuant. La taille de chaque poids est la conductance relative de chaque ion. Dans le cas normal, où trois ions contribuent au potentiel de membrane :
où
- E m est le potentiel de membrane, mesuré en volts
- E X est le potentiel d'équilibre de l'ion X, également en volts
- g X / g tot est la conductance relative de l'ion X, qui est sans dimension
- g tot est la conductance totale de tous les ions perméants en unités arbitraires (par exemple Siemens pour la conductance électrique), dans ce cas g K + + g Na + + g Cl −
Protéines de transport membranaire
Pour la détermination des potentiels de membrane, les deux types les plus importants de protéines de transport d'ions membranaires sont les canaux ioniques et les transporteurs d'ions . Les protéines de canaux ioniques créent des chemins à travers les membranes cellulaires à travers lesquels les ions peuvent diffuser passivement sans dépense directe d'énergie métabolique. Elles ont une sélectivité pour certains ions, ainsi, il existe des canaux ioniques sélectifs pour le potassium , le chlorure et le sodium . Différentes cellules et même différentes parties d'une cellule ( dendrites , corps cellulaires , nœuds de Ranvier ) auront différentes quantités de diverses protéines de transport d'ions. En général, la quantité de certains canaux potassiques est la plus importante pour le contrôle du potentiel de repos (voir ci-dessous). Certaines pompes ioniques telles que la Na+/K+-ATPase sont électrogènes, c'est-à-dire qu'elles produisent un déséquilibre de charge à travers la membrane cellulaire et peuvent également contribuer directement au potentiel de membrane. La plupart des pompes utilisent l'énergie métabolique (ATP) pour fonctionner.
Potentiels d'équilibre
Pour la plupart des cellules animales, les ions potassium (K + ) sont les plus importants pour le potentiel de repos. En raison du transport actif des ions potassium, la concentration de potassium est plus élevée à l'intérieur des cellules qu'à l'extérieur. La plupart des cellules ont des protéines de canaux ioniques sélectifs du potassium qui restent ouvertes en permanence. Il y aura un mouvement net d'ions potassium chargés positivement à travers ces canaux potassiques avec pour résultat une accumulation d'excès de charge négative à l'intérieur de la cellule. Le mouvement vers l'extérieur des ions potassium chargés positivement est dû au mouvement moléculaire aléatoire ( diffusion ) et continue jusqu'à ce qu'une charge négative en excès s'accumule à l'intérieur de la cellule pour former un potentiel membranaire qui peut équilibrer la différence de concentration de potassium entre l'intérieur et l'extérieur de la cellule. "Équilibre" signifie que la force électrique ( potentiel ) qui résulte de l'accumulation de charge ionique , et qui empêche la diffusion vers l'extérieur, augmente jusqu'à ce qu'elle soit égale en amplitude mais opposée en direction à la tendance au mouvement diffusif vers l'extérieur du potassium. Ce point d'équilibre est un potentiel d'équilibre car le flux transmembranaire net (ou courant ) de K + est nul. Une bonne approximation du potentiel d'équilibre d'un ion donné ne nécessite que les concentrations de chaque côté de la membrane et la température. On peut le calculer à l'aide de l' équation de Nernst :
où
- E eq,K + est le potentiel d'équilibre du potassium, mesuré en volts
- R est la constante universelle des gaz , égale à 8,314 joules ·K −1 ·mol −1
- T est la température absolue , mesurée en kelvins (= K = degrés Celsius + 273,15)
- z est le nombre de charges élémentaires de l'ion en question impliquées dans la réaction
- F est la constante de Faraday , égale à 96 485 coulombs ·mol −1 ou J·V −1 ·mol −1
- [K + ] o est la concentration extracellulaire de potassium, mesurée en mol ·m −3 ou mmol·l −1
- [K + ] i est également la concentration intracellulaire de potassium
Les potentiels d'équilibre du potassium d'environ −80 millivolts (à l'intérieur du négatif) sont courants. Des différences sont observées dans différentes espèces, différents tissus au sein du même animal et les mêmes tissus dans différentes conditions environnementales. En appliquant l'équation de Nernst ci-dessus, on peut expliquer ces différences par des changements dans la concentration relative en K + ou des différences de température.
Pour l'usage courant, l'équation de Nernst est souvent donnée sous une forme simplifiée en supposant une température corporelle humaine typique (37 °C), en réduisant les constantes et en passant à Log base 10. (Les unités utilisées pour la concentration ne sont pas importantes car elles s'annulent dans un rapport). Pour le potassium à température corporelle normale, on peut calculer le potentiel d'équilibre en millivolts comme suit :
De même, le potentiel d'équilibre du sodium (Na + ) à la température normale du corps humain est calculé à l'aide de la même constante simplifiée. Vous pouvez calculer E en supposant une concentration extérieure, [K + ] o , de 10 mM et une concentration intérieure, [K + ] i , de 100 mM. Pour les ions chlorure (Cl − ) le signe de la constante doit être inversé (−61,54 mV). Si vous calculez le potentiel d'équilibre du calcium (Ca 2+ ), la charge 2+ divise par deux la constante simplifiée à 30,77 mV. Si vous travaillez à température ambiante, environ 21 °C, les constantes calculées sont d'environ 58 mV pour K + et Na + , −58 mV pour Cl − et 29 mV pour Ca 2+ . À température physiologique, environ 29,5 °C, et à des concentrations physiologiques (qui varient pour chaque ion), les potentiels calculés sont d'environ 67 mV pour Na + , −90 mV pour K + , −86 mV pour Cl − et 123 mV pour Ca 2+ .
Potentiels de repos
Le potentiel de repos de la membrane n'est pas un potentiel d'équilibre car il repose sur une dépense constante d'énergie (pour les pompes ioniques comme mentionné ci-dessus) pour son maintien. Il s'agit d'un potentiel de diffusion dynamique qui prend en compte ce mécanisme, contrairement au potentiel d'équilibre des coussins, ce qui est vrai quelle que soit la nature du système considéré. Le potentiel de repos de la membrane est dominé par l'espèce ionique du système qui a la plus grande conductance à travers la membrane. Pour la plupart des cellules, il s'agit du potassium. Comme le potassium est également l'ion avec le potentiel d'équilibre le plus négatif, le potentiel de repos ne peut généralement pas être plus négatif que le potentiel d'équilibre du potassium. Le potentiel de repos peut être calculé avec l' équation de tension de Goldman-Hodgkin-Katz en utilisant les concentrations d'ions comme pour le potentiel d'équilibre tout en incluant également les perméabilités relatives de chaque espèce ionique. Dans des conditions normales, on peut supposer que seuls les ions potassium, sodium (Na + ) et chlorure (Cl − ) jouent un rôle important pour le potentiel de repos :
Cette équation ressemble à l'équation de Nernst, mais possède un terme pour chaque ion perméant. De plus, z a été inséré dans l'équation, ce qui provoque l' inversion des concentrations intracellulaires et extracellulaires de Cl − par rapport à K + et Na + , car la charge négative du chlorure est gérée en inversant la fraction à l'intérieur du terme logarithmique. * E m est le potentiel de membrane, mesuré en volts * R , T et F sont comme ci-dessus * P s est la perméabilité relative des ions s * [s] Y est la concentration des ions s dans le compartiment Y comme ci-dessus. Une autre façon de visualiser le potentiel de membrane, en considérant plutôt la conductance des canaux ioniques plutôt que la perméabilité de la membrane, est d'utiliser l'équation de Millman (également appelée équation de conductance des cordes) :
ou reformulé
où g tot est la conductance combinée de toutes les espèces ioniques, encore une fois en unités arbitraires. Cette dernière équation décrit le potentiel de membrane au repos comme une moyenne pondérée des potentiels d'inversion du système, où les poids sont les conductances relatives de chaque espèce d'ion ( g X / g tot ). Pendant le potentiel d'action, ces poids changent. Si les conductances de Na + et Cl − sont nulles, le potentiel de membrane se réduit au potentiel de Nernst pour K + (comme g K + = g tot ). Normalement, dans des conditions de repos g Na+ et g Cl− ne sont pas nuls, mais ils sont bien plus petits que g K+ , ce qui rend E m proche de E eq,K+ . Les états médicaux tels que l'hyperkaliémie dans lesquels le potassium sérique sanguin (qui gouverne [K + ] o ) est modifié sont très dangereux car ils compensent E eq,K+ , affectant ainsi E m . Cela peut provoquer des arythmies et un arrêt cardiaque . L'utilisation d'une injection en bolus de chlorure de potassium dans les exécutions par injection létale arrête le cœur en décalant le potentiel de repos vers une valeur plus positive, ce qui dépolarise et contracte les cellules cardiaques de façon permanente, ne permettant pas au cœur de se repolariser et ainsi d'entrer en diastole pour se remplir de sang.
Bien que l'équation de tension GHK et l'équation de Millman soient liées, elles ne sont pas équivalentes. La différence essentielle est que l'équation de Millman suppose que la relation courant-tension est ohmique, tandis que l'équation de tension GHK prend en compte les petites rectifications instantanées prédites par l' équation de flux GHK causées par le gradient de concentration des ions. Ainsi, une estimation plus précise du potentiel de membrane peut être calculée en utilisant l'équation GHK qu'avec l'équation de Millman.
Mesure des potentiels de repos
Dans certaines cellules, le potentiel de membrane change constamment (comme dans les cellules du stimulateur cardiaque ). Pour ces cellules, il n'y a jamais de « repos » et le « potentiel de repos » est un concept théorique. D'autres cellules, dont les fonctions de transport membranaire sont peu nombreuses et qui changent avec le temps, ont un potentiel de membrane de repos qui peut être mesuré en insérant une électrode dans la cellule. Les potentiels transmembranaires peuvent également être mesurés optiquement à l'aide de colorants qui modifient leurs propriétés optiques en fonction du potentiel de membrane.
Résumé des valeurs du potentiel de repos dans différents types de cellules
Histoire
Les courants de repos dans les nerfs ont été mesurés et décrits par Julius Bernstein en 1902, où il a proposé une « théorie de la membrane » qui expliquait le potentiel de repos des nerfs et des muscles comme un potentiel de diffusion.