Ardipithecus est un genre d' hominidé éteint qui a vécu à la fin du Miocène et au début du Pliocène dans la dépression d'Afar , en Éthiopie . Décrit à l'origine comme l'un des premiers ancêtres de l'homme après sa divergence avec les chimpanzés, la relation de ce genre avec les ancêtres humains et la question de savoir s'il s'agit d'un hominidé sont aujourd'hui sujettes à débat.espèces fossilessont décrites dans la littérature : A. ramidus , qui a vécu il y a environ 4,4 millions d'années au début du Pliocène , et A. kadabba , daté d'environ 5,6 millions d'années (fin du Miocène ). Une analyse comportementale initiale a indiqué qu'Ardipithecus pourrait être très similaire aux chimpanzés, cependant une analyse plus récente basée sur la taille des canines et l'absence de dimorphisme sexuel canin indique qu'Ardipithecus était caractérisé par une agressivité réduite, et qu'ils ressemblent davantage aux bonobos .
Certaines analyses décrivent l'Australopithèque comme étant spécifiquement le frère d' Ardipithecus ramidus . Cela signifie que l'Australopithèque est nettement plus étroitement lié à Ardipithecus ramidus qu'à Ardipithecus kadabba . D' un point de vue cladistique , l'Australopithèque (et finalement l'Homo sapiens ) a effectivement émergé au sein de la lignée d'Ardipithecus , et cette lignée n'est pas littéralement éteinte.
Ardipithecus ramidus
Ardipithecus ramidus a été nommé en septembre 1994. Le premier fossile découvert a été daté d'il y a 4,4 millions d'années sur la base de sa position stratigraphique entre deux couches volcaniques : le complexe de tuf basal de Gaala (GATC) et le tuf basaltique de Daam Aatu (DABT). Le nom Ardipithecus ramidus provient principalement de la langue afar , dans laquelle Ardi signifie « sol/étage » et ramid signifie « racine ». La partie pithecus du nom provient du mot grec pour « singe ».
Comme la plupart des hominidés , mais contrairement à tous les hominidés précédemment reconnus, il avait un hallux ou gros orteil de préhension adapté à la locomotion dans les arbres. On ne sait pas combien d'autres caractéristiques de son squelette reflètent également l'adaptation à la bipédie au sol. Comme les hominidés ultérieurs, Ardipithecus avait des canines réduites et un dimorphisme sexuel canin réduit.
En 1992-1993, une équipe de recherche dirigée par Tim White a découvert les premiers fossiles d'A. ramidus - dix-sept fragments comprenant un crâne, une mandibule, des dents et des os de bras - dans la dépression d'Afar , dans la vallée moyenne de la rivière Awash en Éthiopie . D'autres fragments ont été récupérés en 1994, représentant 45 % du squelette total. Ce fossile a été décrit à l'origine comme une espèce d' Australopithecus , mais White et ses collègues ont ensuite publié une note dans la même revue renommant le fossile sous un nouveau genre, Ardipithecus . Entre 1999 et 2003, une équipe multidisciplinaire dirigée par Sileshi Semaw a découvert des os et des dents de neuf individus d'A. ramidus à As Duma, dans la région de Gona , dans la région d'Afar en Éthiopie . Les fossiles ont été datés d'un âge compris entre 4,35 et 4,45 millions d'années.
.svg/1280px-Map_of_the_fossil_sites_of_the_earliest_hominids_(35.8-3.3M_BP).svg.png)
Ardipithecus ramidus avait un petit cerveau, mesurant entre 300 et 350 cm3 . Il est légèrement plus petit que le cerveau d'un bonobo ou d'un chimpanzé femelle moderne, mais beaucoup plus petit que celui d' australopithèques comme Lucy (~400 à 550 cm3 ) et environ 20 % de la taille du cerveau d'Homo sapiens moderne . Comme les chimpanzés communs, A. ramidus était beaucoup plus prognathe que les humains modernes.
Les dents d' A. ramidus n'avaient pas la spécialisation des autres singes et suggèrent qu'il s'agissait d'un omnivore et d'un frugivore généralisé (mangeur de fruits) dont le régime alimentaire ne dépendait pas fortement du feuillage, des matières végétales fibreuses (racines, tubercules, etc.) ou d'aliments durs et/ou abrasifs. La taille de la canine supérieure des mâles d' A. ramidus n'était pas nettement différente de celle des femelles. Leurs canines supérieures étaient moins acérées que celles des chimpanzés communs modernes en partie à cause de cette taille réduite de la canine supérieure, car les canines supérieures plus grandes peuvent être aiguisées par l'usure contre les dents de la bouche inférieure. Les caractéristiques de la canine supérieure d' A. ramidus contrastent avec le dimorphisme sexuel observé chez les chimpanzés communs, où les mâles ont des canines supérieures nettement plus grandes et plus acérées que les femelles. Parmi les singes vivants, les bonobos ont le plus petit dimorphisme sexuel canin, bien qu'il soit toujours plus grand que celui affiché par A. ramidus .
La nature moins prononcée des canines supérieures chez A. ramidus a été utilisée pour déduire certains aspects du comportement social de l'espèce et d'hominidés plus ancestraux. En particulier, elle a été utilisée pour suggérer que le dernier ancêtre commun des hominidés et des singes africains était caractérisé par relativement peu d'agressivité entre mâles et entre groupes. Cela est nettement différent des modèles sociaux des chimpanzés communs, chez lesquels l'agressivité entre mâles et entre groupes est généralement élevée. Les chercheurs d'une étude de 2009 ont déclaré que cette condition « compromet le chimpanzé vivant en tant que modèle comportemental pour la condition d'hominidé ancestral ». La taille des canines et le dimorphisme sexuel des canines du bonobo ressemblent davantage à ceux d' A. ramidus , et par conséquent, les bonobos sont désormais suggérés comme modèle comportemental.
A. ramidus a existé plus récemment que l' ancêtre commun le plus récent des humains et des chimpanzés ( CLCA ou Pan - Homo LCA ) et n'est donc pas entièrement représentatif de cet ancêtre commun. Néanmoins, il est à certains égards différent des chimpanzés, ce qui suggère que l'ancêtre commun diffère du chimpanzé moderne. Après la divergence des lignées des chimpanzés et des humains, les deux ont subi des changements évolutifs substantiels. Les pieds des chimpanzés sont spécialisés pour saisir les arbres ; les pieds d'A. ramidus sont mieux adaptés à la marche. Les canines d' A. ramidus sont plus petites et de taille égale entre les mâles et les femelles, ce qui suggère une réduction des conflits entre mâles, une augmentation des liens de couple et un investissement parental accru. « Ainsi, des changements fondamentaux dans le comportement reproductif et social se sont probablement produits chez les hominidés bien avant qu'ils aient un cerveau élargi et commencent à utiliser des outils en pierre », a conclu l'équipe de recherche.
Ardi
Le 1er octobre 2009 , les paléontologues ont officiellement annoncé la découverte du squelette fossile relativement complet d'A. ramidus, découvert pour la première fois en 1994. Le fossile est le reste d'une femelle de 50 kg au cerveau de petite taille, surnommée « Ardi », et comprend la majeure partie du crâne et des dents, ainsi que le bassin, les mains et les pieds. le désert d'Afar en Éthiopie, sur un site appelé Aramis, dans la région du Moyen-Awash. La datation radiométrique des couches de cendres volcaniques recouvrant les dépôts suggère qu'Ardi a vécu il y a environ 4,3 à 4,5 millions d'années. Cette date a cependant été remise en question par d'autres. Fleagle et Kappelman suggèrent que la région dans laquelle Ardi a été trouvé est difficile à dater radiométriquement, et ils soutiennent qu'Ardi devrait être daté à 3,9 millions d'années.
Le fossile est considéré par ses descripteurs comme un révélateur d'une étape de l'évolution humaine dont on savait peu de choses, plus d'un million d'années avant Lucy ( Australopithecus afarensis ), l'ancêtre emblématique de l'homme primitif qui a vécu il y a 3,2 millions d'années et a été découvert en 1974 à seulement 74 km du site de découverte d'Ardi. Cependant, comme le squelette d'« Ardi » n'est pas plus vieux de 200 000 ans que les premiers fossiles d' Australopithecus , et pourrait en fait être plus jeune qu'eux, certains chercheurs doutent qu'il puisse représenter un ancêtre direct d' Australopithecus .
Certains chercheurs déduisent de la forme de son bassin et de ses membres et de la présence de son hallux abductif , que « Ardi » était un bipède facultatif : bipède lorsqu'il se déplaçait sur le sol, mais quadrupède lorsqu'il se déplaçait dans les branches des arbres. A. ramidus avait une capacité de marche plus primitive que les hominidés ultérieurs et ne pouvait pas marcher ou courir sur de longues distances. Les dents suggèrent l'omnivorie et sont plus généralisées que celles des singes modernes.
-
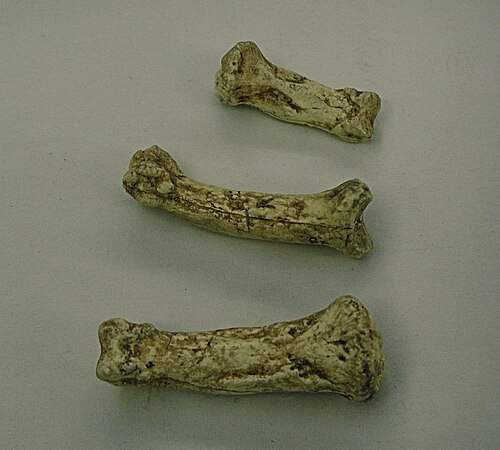 Moulages des os des doigts d'Ardi.
Moulages des os des doigts d'Ardi.
.jpg/1280px-Ardipithecus_(finger_bones).jpg)
Ardipithèque kadabba

Ardipithecus kadabba est « connu uniquement par des dents et des morceaux d'os squelettiques » et est daté d'environ 5,6 millions d'années . Il a été décrit comme une « chronoespèce probable » (c'est-à-dire un ancêtre) d' A. ramidus . Bien qu'il ait été initialement considéré comme une sous-espèce d' A. ramidus , en 2004, les anthropologues Yohannes Haile-Selassie , Gen Suwa et Tim D. White ont publié un article élevant A. kadabba au niveau d'espèce sur la base de dents récemment découvertes en Éthiopie . Ces dents présentent une « morphologie primitive et un modèle d'usure » qui démontrent qu'A . kadabba est une espèce distincte d' A. ramidus .
Le nom spécifique vient du mot Afar qui signifie « ancêtre familial de base ».
Classification
Français En raison de plusieurs caractéristiques communes avec les chimpanzés, de sa proximité avec la période de divergence des singes et de son incomplétude fossile, la position exacte d' Ardipithecus dans le registre fossile est un sujet de controverse. Le primatologue Esteban Sarmiento a systématiquement comparé et conclu qu'il n'y avait pas suffisamment de preuves anatomiques pour soutenir une lignée exclusivement humaine. Sarmiento a noté qu'Ardipithecus ne partage aucune caractéristique exclusive aux humains, et certaines de ses caractéristiques (celles du poignet et du basicrâne) suggèrent qu'il a divergé des humains avant le dernier ancêtre commun homme-gorille . Son étude comparative ( allométrie étroite ) en 2011 sur les longueurs des molaires et des segments corporels (qui incluaient des primates vivants de taille corporelle similaire) a noté que certaines dimensions, notamment les membres supérieurs courts et les métacarpes , rappellent celles des humains, mais d'autres dimensions telles que les longs orteils et la surface molaire relative sont semblables à celles des grands singes . Sarmiento a conclu que de telles mesures de longueur peuvent changer dans les deux sens au cours de l'évolution et ne sont pas de très bons indicateurs de parenté ( homoplasie ).
Cependant, certaines études ultérieures plaident toujours en faveur de sa classification dans la lignée humaine. En 2014, il a été signalé que les os de la main d' Ardipithecus , d'Australopithecus sediba et d'A. afarensis présentent le troisième processus styloïde métacarpien , qui est absent chez d'autres singes. Des organisations cérébrales uniques (telles que le déplacement latéral des foramens carotidiens, l'abréviation médiolatérale du tympan latéral et un élément basioccipital trapézoïdal raccourci ) chez Ardipithecus ne se retrouvent également que chez les Australopithecus et Homo . La comparaison de la morphologie des racines dentaires avec celles du Sahelanthropus antérieur a également indiqué une forte ressemblance, indiquant également une inclusion dans la lignée humaine.
Arbre évolutif selon une étude de 2019 :