Les lymphocytes B , également appelés lymphocytes B , sont un type de globule blanc du sous-type lymphocyte . Ils fonctionnent dans la composante de l'immunité humorale du système immunitaire adaptatif . Les lymphocytes B produisent des molécules d'anticorps qui peuvent être sécrétées ou insérées dans la membrane plasmique où elles font partie des récepteurs des lymphocytes B. Lorsqu'un lymphocyte B naïf ou mémoire est activé par un antigène, il prolifère et se différencie en une cellule effectrice sécrétant des anticorps , appelée plasmablaste ou plasmocyte. De plus, les lymphocytes B présentent des antigènes (ils sont également classés comme cellules présentatrices d'antigènes professionnelles, APC ) et sécrètent des cytokines . Chez les mammifères, les lymphocytes B mûrissent dans la moelle osseuse , qui se trouve au cœur de la plupart des os . Chez les oiseaux , les cellules B mûrissent dans la bourse de Fabricius , un organe lymphoïde où elles ont été découvertes pour la première fois par Chang et Glick, c'est pourquoi le B signifie bourse et non moelle osseuse , comme on le croit généralement.
Les cellules B, contrairement aux deux autres classes de lymphocytes, les cellules T et les cellules tueuses naturelles , expriment des récepteurs de cellules B (BCR) sur leur membrane cellulaire . Les BCR permettent à la cellule B de se lier à un antigène étranger , contre lequel elle initiera une réponse anticorps. Les récepteurs des cellules B sont extrêmement spécifiques, tous les BCR d'une cellule B reconnaissant le même épitope .
Développement
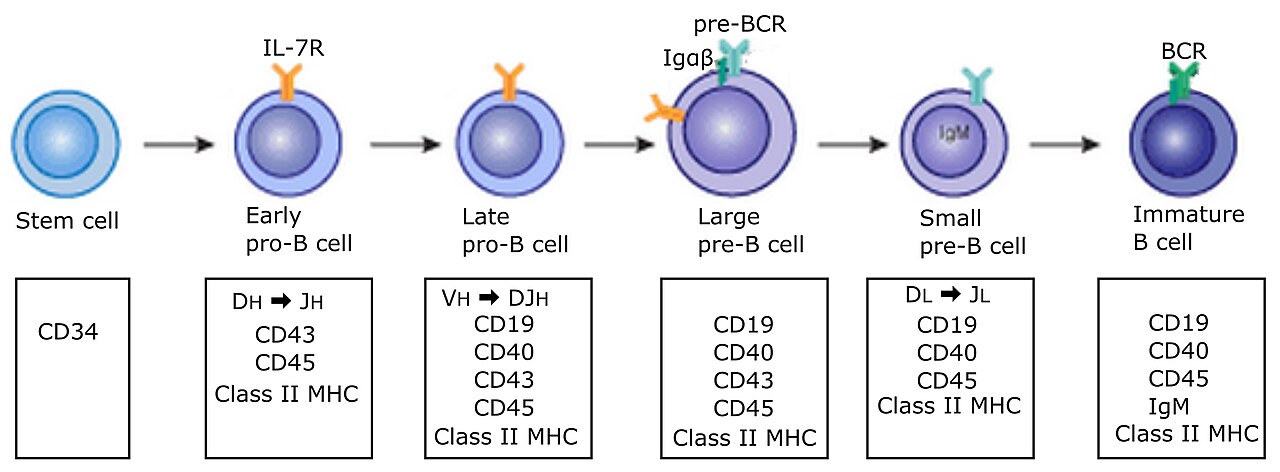
Les cellules B se développent à partir de cellules souches hématopoïétiques (HSC) qui proviennent de la moelle osseuse . Les HSC se différencient d'abord en cellules progénitrices multipotentes (MPP), puis en cellules progénitrices lymphoïdes communes (CLP). À partir de là, leur développement en cellules B se déroule en plusieurs étapes (illustrées dans l'image de droite), chacune marquée par divers modèles d'expression génétique et des arrangements de locus de gènes de chaîne H et de chaîne L des immunoglobulines , ces derniers étant dus aux cellules B subissant une recombinaison V(D)J au cours de leur développement.
Les cellules B subissent deux types de sélection lors de leur développement dans la moelle osseuse pour assurer un développement approprié, tous deux impliquant des récepteurs des cellules B (BCR) à la surface de la cellule. La sélection positive se produit par une signalisation indépendante de l'antigène impliquant à la fois le pré-BCR et le BCR. Si ces récepteurs ne se lient pas à leur ligand , les cellules B ne reçoivent pas les signaux appropriés et cessent de se développer. La sélection négative se produit par la liaison de l'auto-antigène avec le BCR ; si le BCR peut se lier fortement à l'auto-antigène, alors la cellule B subit l'un des quatre destins suivants : suppression clonale , édition du récepteur , anergie ou ignorance (la cellule B ignore le signal et continue son développement). Ce processus de sélection négative conduit à un état de tolérance centrale , dans lequel les cellules B matures ne se lient pas aux auto-antigènes présents dans la moelle osseuse.
Pour terminer leur développement, les cellules B immatures migrent de la moelle osseuse vers la rate en tant que cellules B de transition , en passant par deux stades de transition : T1 et T2. Tout au long de leur migration vers la rate et après leur entrée dans la rate, elles sont considérées comme des cellules B T1. Dans la rate, les cellules B T1 se transforment en cellules B T2. Les cellules B T2 se différencient en cellules B folliculaires (FO) ou en cellules B de la zone marginale (MZ) en fonction des signaux reçus via le BCR et d'autres récepteurs. Une fois différenciées, elles sont désormais considérées comme des cellules B matures ou des cellules B naïves.
Activation

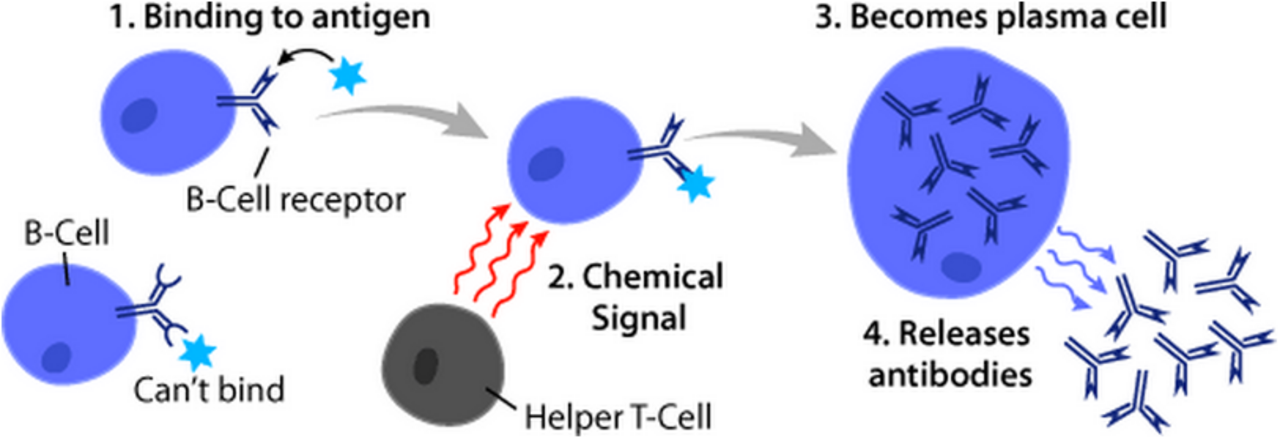
L'activation des lymphocytes B se produit dans les organes lymphoïdes secondaires (SLO), tels que la rate et les ganglions lymphatiques . Une fois que les lymphocytes B ont mûri dans la moelle osseuse, ils migrent à travers le sang vers les SLO, qui reçoivent un apport constant d'antigène par la lymphe circulante . Au SLO, l'activation des lymphocytes B commence lorsque le lymphocyte B se lie à un antigène via son BCR. Bien que les événements qui se produisent immédiatement après l'activation n'aient pas encore été complètement déterminés, on pense que les lymphocytes B sont activés conformément au modèle de ségrégation cinétique , initialement déterminé dans les lymphocytes T. Ce modèle indique qu'avant la stimulation antigénique, les récepteurs diffusent à travers la membrane entrant en contact avec Lck et CD45 à fréquence égale, créant un équilibre net de phosphorylation et de non-phosphorylation. Ce n'est que lorsque la cellule entre en contact avec une cellule présentatrice d'antigène que le CD45 plus gros est déplacé en raison de la distance étroite entre les deux membranes. Cela permet la phosphorylation nette du BCR et l'initiation de la voie de transduction du signal . Parmi les trois sous-ensembles de cellules B, les cellules B FO subissent préférentiellement une activation dépendante des cellules T tandis que les cellules B MZ et les cellules B B1 subissent préférentiellement une activation indépendante des cellules T.
L'activation des cellules B est renforcée par l'activité de CD21 , un récepteur de surface en complexe avec les protéines de surface CD19 et CD81 (les trois sont collectivement connues sous le nom de complexe corécepteur des cellules B). Lorsqu'un BCR se lie à un antigène marqué par un fragment de la protéine du complément C3, CD21 se lie au fragment C3, se lie au BCR lié et les signaux sont transduits via CD19 et CD81 pour abaisser le seuil d'activation de la cellule.
Activation dépendante des cellules T
Les antigènes qui activent les cellules B avec l'aide des cellules T sont connus sous le nom d'antigènes dépendants des cellules T (TD) et comprennent des protéines étrangères. Ils sont nommés ainsi parce qu'ils sont incapables d'induire une réponse humorale dans les organismes dépourvus de cellules T. Les réponses des cellules B à ces antigènes prennent plusieurs jours, bien que les anticorps générés aient une affinité plus élevée et soient plus polyvalents sur le plan fonctionnel que ceux générés par une activation indépendante des cellules T.
Français Une fois qu'un BCR se lie à un antigène TD, l'antigène est absorbé dans la cellule B par endocytose médiée par un récepteur , dégradé et présenté aux cellules T sous forme de morceaux de peptides en complexe avec des molécules du CMH-II sur la membrane cellulaire. T auxiliaires (T H ) , généralement les cellules T auxiliaires folliculaires (TFH ) reconnaissent et se lient à ces complexes CMH-II-peptide par l'intermédiaire de leur récepteur des cellules T (TCR) . Après la liaison TCR-CMH-II-peptide, les cellules T expriment la protéine de surface CD40L ainsi que des cytokines telles que l'IL-4 et l'IL-21 . CD40L sert de facteur de co-stimulation nécessaire à l'activation des cellules B en se liant au récepteur de surface des cellules B CD40 , qui favorise la prolifération des cellules B , le changement de classe d'immunoglobuline et l'hypermutation somatique ainsi que soutient la croissance et la différenciation des cellules T. Les cytokines dérivées des cellules T liées aux récepteurs des cytokines des cellules B favorisent également la prolifération des cellules B, le changement de classe d'immunoglobuline et l'hypermutation somatique, ainsi que la différenciation guidée. Une fois que les cellules B ont reçu ces signaux, elles sont considérées comme activées.
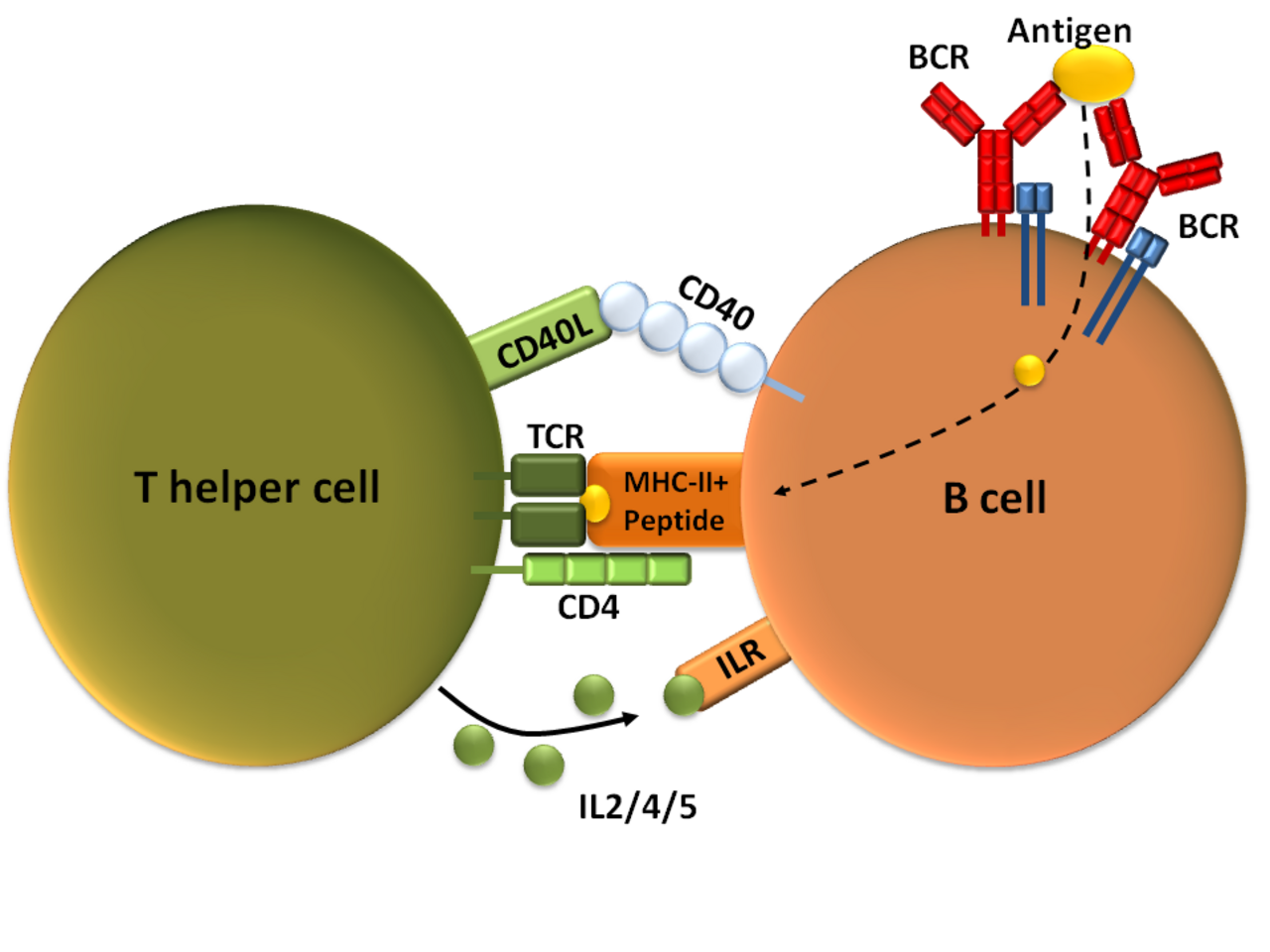
Une fois activées, les cellules B participent à un processus de différenciation en deux étapes qui produit à la fois des plasmoblastes à courte durée de vie pour une protection immédiate et des plasmocytes à longue durée de vie et des cellules B à mémoire pour une protection persistante. La première étape, connue sous le nom de réponse extrafolliculaire, se produit à l'extérieur des follicules lymphoïdes mais toujours dans le SLO. Au cours de cette étape, les cellules B activées prolifèrent, peuvent subir un changement de classe d'immunoglobuline et se différencier en plasmoblastes qui produisent des anticorps précoces et faibles, principalement de classe IgM.

La deuxième étape consiste en l'entrée des cellules B activées dans un follicule lymphoïde et la formation d'un centre germinatif (GC) , qui est un microenvironnement spécialisé dans lequel les cellules B subissent une prolifération extensive, une commutation de classe d'immunoglobuline et une maturation d'affinité dirigée par une hypermutation somatique. Ces processus sont facilités par T FH et les cellules dendritiques folliculaires au sein du GC et génèrent à la fois des cellules B mémoires à haute affinité et des cellules plasmatiques à longue durée de vie. Les cellules plasmatiques résultantes sécrètent un grand nombre d'anticorps et restent dans le SLO ou, plus préférentiellement, migrent vers la moelle osseuse.
Activation indépendante des cellules T
Les antigènes qui activent les cellules B sans l'aide des cellules T sont appelés antigènes indépendants des cellules T (TI) et comprennent des polysaccharides étrangers et de l'ADN CpG non méthylé. Ils sont nommés ainsi parce qu'ils sont capables d'induire une réponse humorale dans les organismes dépourvus de cellules T. La réponse des cellules B à ces antigènes est rapide, bien que les anticorps générés aient tendance à avoir une affinité plus faible et soient moins polyvalents sur le plan fonctionnel que ceux générés par l'activation dépendante des cellules T.
Comme pour les antigènes TD, les cellules B activées par les antigènes TI ont besoin de signaux supplémentaires pour terminer l'activation, mais au lieu de les recevoir des cellules T, ils sont fournis soit par la reconnaissance et la liaison d'un constituant microbien commun aux récepteurs de type Toll (TLR) , soit par une réticulation étendue des BCR à des épitopes répétés sur une cellule bactérienne. Les cellules B activées par les antigènes TI continuent à proliférer à l'extérieur des follicules lymphoïdes mais toujours dans les SLO (les GC ne se forment pas), subissent éventuellement un changement de classe d'immunoglobuline et se différencient en plasmablastes de courte durée qui produisent des anticorps précoces et faibles, principalement de classe IgM, mais aussi certaines populations de plasmocytes à longue durée de vie.
Activation des cellules B à mémoire
L'activation des lymphocytes B mémoires commence par la détection et la liaison de leur antigène cible, qui est partagé par leur lymphocyte B parent. Certains lymphocytes B mémoires peuvent être activés sans l'aide des lymphocytes T, comme certains lymphocytes B mémoires spécifiques des virus, mais d'autres ont besoin de l'aide des lymphocytes T. Lors de la liaison à l'antigène, le lymphocyte B mémoire absorbe l'antigène par endocytose à médiation par récepteur, le dégrade et le présente aux lymphocytes T sous forme de morceaux de peptides en complexe avec les molécules du CMH-II sur la membrane cellulaire. Les lymphocytes T auxiliaires mémoire (T H ), généralement les lymphocytes T auxiliaires folliculaires mémoire (T FH ), qui sont dérivés de lymphocytes T activés avec le même antigène, reconnaissent et lient ces complexes CMH-II-peptide par l'intermédiaire de leur TCR. Suite à la liaison du peptide TCR-MHC-II et au relais d'autres signaux provenant de la cellule mémoire T FH , la cellule B mémoire est activée et se différencie soit en plasmablastes et en plasmocytes via une réponse extrafolliculaire, soit entre dans une réaction du centre germinatif où elle génère des plasmocytes et davantage de cellules B mémoire. On ne sait pas si les cellules B mémoire subissent une maturation d'affinité supplémentaire au sein de ces GC secondaires. in vitro des cellules B mémoire peut être obtenue par stimulation avec divers activateurs, tels que le mitogène du phytolaque ou les anticorps monoclonaux anti- CD40 , cependant, une étude a révélé qu'une combinaison de R-848 et d'IL-2 humaine recombinante était l'activateur le plus efficace.
Types de cellules B
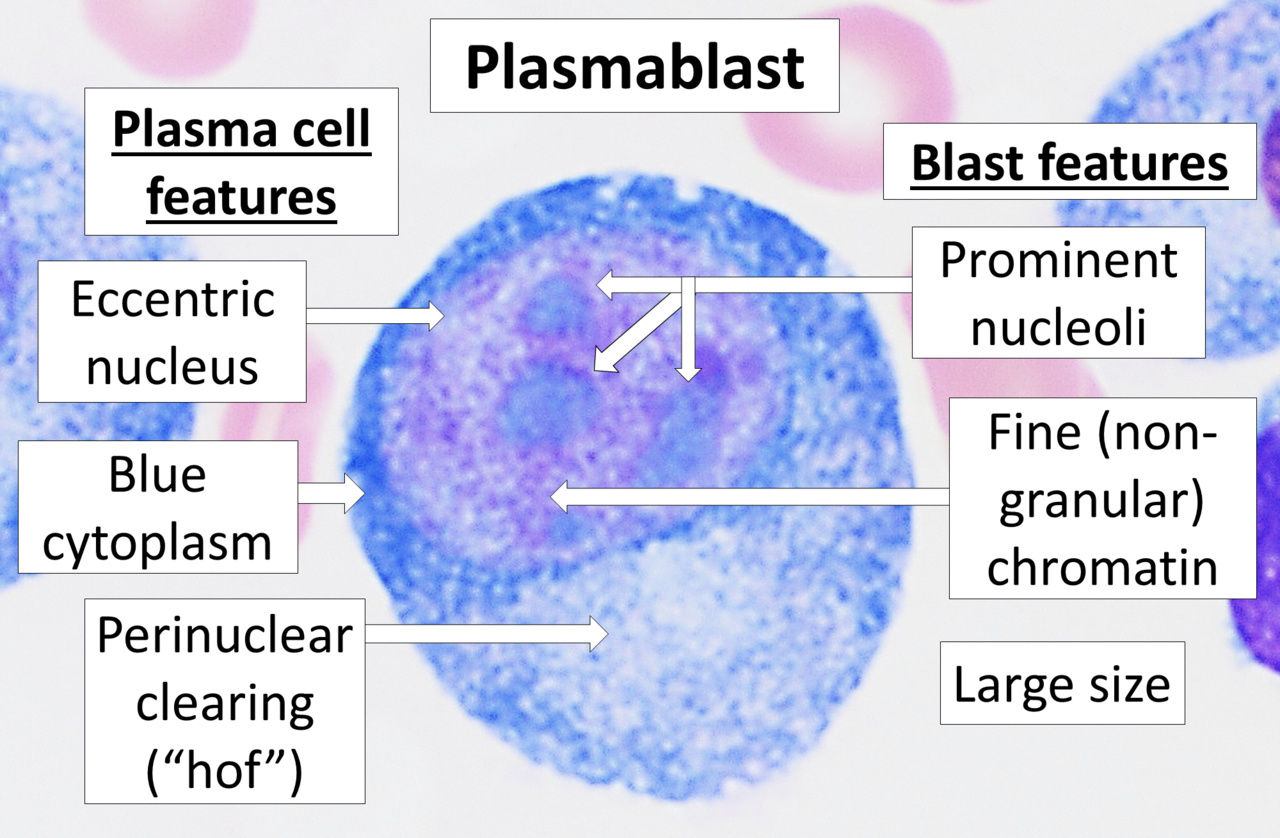
- Plasmablaste
- Une cellule sécrétant des anticorps à courte durée de vie et proliférante, issue de la différenciation des cellules B. Les plasmoblastes sont générés au début d'une infection et leurs anticorps ont tendance à avoir une affinité plus faible envers leur antigène cible par rapport aux plasmocytes. Les plasmoblastes peuvent résulter de l'activation indépendante des cellules T des cellules B ou de la réponse extrafolliculaire de l'activation dépendante des cellules T des cellules B.
- Cellule plasmatique
- Cellule sécrétant des anticorps non proliférante et à longue durée de vie issue de la différenciation des cellules B. Il existe des preuves que les cellules B se différencient d'abord en une cellule de type plasmoblaste, puis en une cellule plasmatique. Les cellules plasmatiques sont générées plus tard au cours d'une infection et, par rapport aux plasmablastes, ont des anticorps avec une affinité plus élevée envers leur antigène cible en raison de la maturation d'affinité dans le centre germinatif (GC) et produisent plus d'anticorps. Les cellules plasmatiques résultent généralement de la réaction du centre germinatif de l'activation des cellules B dépendante des cellules T, bien qu'elles puissent également résulter de l'activation des cellules B indépendante des cellules T.
- Cellule lymphoplasmacytoïde
- Cellule présentant un mélange de caractéristiques morphologiques de lymphocytes B et de plasmocytes, que l'on pense être étroitement liée aux plasmocytes ou en être un sous-type. Ce type de cellule se retrouve dans les dyscrasies plasmocytaires pré-malignes et malignes associées à la sécrétion de protéines monoclonales IgM ; ces dyscrasies comprennent la gammapathie monoclonale IgM de signification indéterminée et la macroglobulinémie de Waldenström .
- Cellule B à mémoire
- Cellule B dormante issue de la différenciation des cellules B. Leur fonction est de circuler dans le corps et d'initier une réponse anticorps plus forte et plus rapide (connue sous le nom de réponse anticorps secondaire anamnestique) si elles détectent l'antigène qui a activé leur cellule B parente (les cellules B mémoire et leurs cellules B parentes partagent le même BCR, elles détectent donc le même antigène). Les cellules B mémoire peuvent être générées à partir d'une activation dépendante des cellules T par la réponse extrafolliculaire et la réaction du centre germinatif ainsi que par l'activation indépendante des cellules T des cellules B1.
- Cellule B-2
- Cellules B FO et cellules B MZ.
- Cellule B folliculaire (FO) (également appelée cellule B-2)
- Le type de lymphocytes B le plus courant et, lorsqu'il ne circule pas dans le sang, on le trouve principalement dans les follicules lymphoïdes des organes lymphoïdes secondaires (OLS). Ils sont responsables de la génération de la majorité des anticorps à haute affinité lors d'une infection.
- Cellule B de la zone marginale (MZ)
- On les trouve principalement dans la zone marginale de la rate et ils servent de première ligne de défense contre les agents pathogènes transmissibles par le sang, car la zone marginale reçoit de grandes quantités de sang de la circulation générale. Ils peuvent subir une activation indépendante et dépendante des cellules T, mais subissent préférentiellement une activation indépendante des cellules T.
- Cellule B-1
- Elles proviennent d'une voie de développement différente des cellules B FO et des cellules B MZ. Chez la souris, elles peuplent principalement la cavité péritonéale et la cavité pleurale , génèrent des anticorps naturels (anticorps produits sans infection), se défendent contre les agents pathogènes des muqueuses et présentent principalement une activation indépendante des cellules T. Un véritable homologue des cellules B-1 de souris n'a pas été découvert chez l'homme, bien que diverses populations cellulaires similaires aux cellules B-1 aient été décrites.
- Cellule B régulatrice (Breg)
- Un type de cellule B immunosuppressive qui arrête l'expansion des lymphocytes pathogènes pro-inflammatoires par la sécrétion d'IL-10, d'IL-35 et de TGF-β. De plus, il favorise la génération de cellules T régulatrices (Treg) en interagissant directement avec les cellules T pour orienter leur différenciation vers les Treg. Aucune identité commune de cellule Breg n'a été décrite et de nombreux sous-ensembles de cellules Breg partageant des fonctions régulatrices ont été trouvés chez la souris et l'homme. On ne sait pas actuellement si les sous-ensembles de cellules Breg sont liés au développement et comment se produit exactement la différenciation en une cellule Breg. Il existe des preuves montrant que presque tous les types de cellules B peuvent se différencier en une cellule Breg par des mécanismes impliquant des signaux inflammatoires et la reconnaissance BCR.
Pathologie liée aux cellules B
Les maladies auto-immunes peuvent résulter d'une reconnaissance anormale des auto-antigènes par les cellules B suivie de la production d'auto-anticorps. Les maladies auto-immunes où l'activité de la maladie est corrélée à l'activité des cellules B comprennent la sclérodermie , la sclérose en plaques , le lupus érythémateux disséminé , le diabète de type 1 , le syndrome du côlon irritable post-infectieux et la polyarthrite rhumatoïde .
La transformation maligne des cellules B et de leurs précurseurs peut provoquer une multitude de cancers , notamment la leucémie lymphoïde chronique (LLC) , la leucémie lymphoblastique aiguë (LAL) , la leucémie à tricholeucocytes , le lymphome folliculaire , le lymphome non hodgkinien , le lymphome de Hodgkin et les tumeurs malignes des plasmocytes telles que le myélome multiple , la macroglobulinémie de Waldenström et certaines formes d' amylose .
Les cellules B anormales peuvent être relativement grandes et certaines maladies incluent cela dans leur nom, comme les lymphomes diffus à grandes cellules B (LDGCB) et le lymphome intravasculaire à grandes cellules B.
Les patients atteints d’alymphocytose à cellules B sont prédisposés aux infections.
Épigénétique
Une étude qui a examiné le méthylome des cellules B tout au long de leur cycle de différenciation, en utilisant le séquençage bisulfite du génome entier (WGBS), a montré qu'il existe une hypométhylation des stades les plus précoces aux stades les plus différenciés. La plus grande différence de méthylation se situe entre les stades des cellules B du centre germinatif et les cellules B mémoires. De plus, cette étude a montré qu'il existe une similitude entre les tumeurs des cellules B et les cellules B à longue durée de vie dans leurs signatures de méthylation de l'ADN .