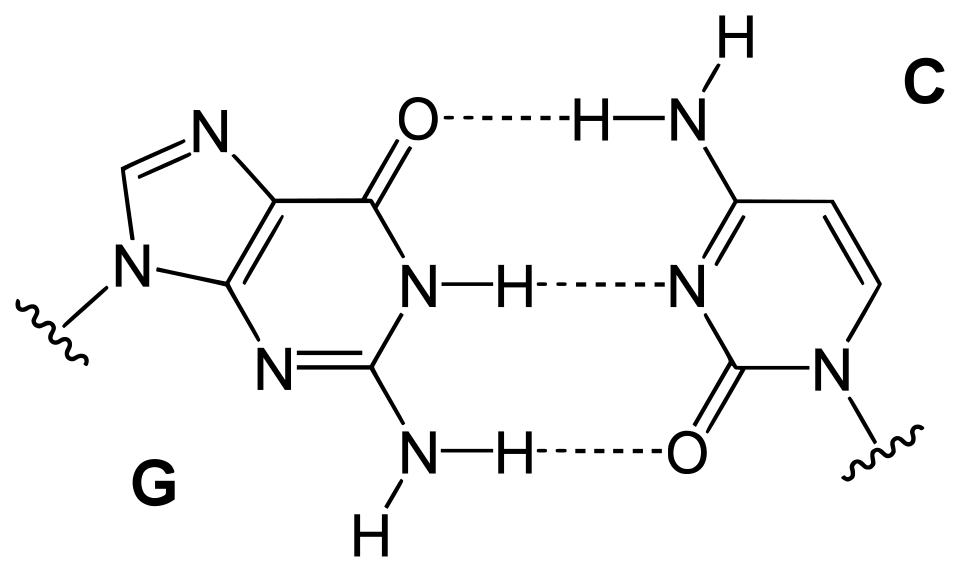
Une paire de bases ( pb ) est l'unité fondamentale des acides nucléiques double brin, constituée de deux nucléobases liées par des liaisons hydrogène . Elles forment les éléments constitutifs de la double hélice d'ADN et contribuent à la structure repliée de l'ADN et de l'ARN . Déterminées par des motifs de liaisons hydrogène spécifiques , les paires de bases « Watson - Crick » (ou « Watson-Crick- Franklin ») ( guanine - cytosine et adénine - thymine / uracile ) permettent à l'hélice d'ADN de conserver une structure hélicoïdale régulière, subtilement dépendante de sa séquence nucléotidique . La complémentarité de cette structure appariée à base fournit une copie redondante de l' information génétique codée dans chaque brin d'ADN. La structure régulière et la redondance des données offertes par la double hélice d'ADN rendent ce dernier particulièrement adapté au stockage de l'information génétique. L'appariement des bases entre l'ADN et les nucléotides entrants constitue le mécanisme par lequel l'ADN polymérase réplique l'ADN et l' ARN polymérase le transcrit en ARN. De nombreuses protéines de liaison à l'ADN peuvent reconnaître des motifs d'appariement de bases spécifiques qui identifient des régions régulatrices particulières des gènes.
Les paires de bases intramoléculaires peuvent se former au sein des acides nucléiques simple brin. Ceci est particulièrement important dans les molécules d'ARN (par exemple, l'ARN de transfert ), où les paires de bases Watson-Crick (guanine-cytosine et adénine-uracile) permettent la formation de courtes hélices double brin, et une grande variété d'interactions non-Watson-Crick (par exemple, G-U ou A-A) permettent aux ARN de se replier en une vaste gamme de structures tridimensionnelles spécifiques . De plus, l'appariement des bases entre l'ARN de transfert (ARNt) et l'ARN messager (ARNm) constitue la base des événements de reconnaissance moléculaire qui aboutissent à la traduction de la séquence nucléotidique de l'ARNm en séquence d'acides aminés des protéines via le code génétique .
La taille d'un gène individuel ou du génome entier d'un organisme est souvent mesurée en paires de bases, car l'ADN est généralement double brin. Ainsi, le nombre total de paires de bases est égal au nombre de nucléotides dans l'un des brins (à l'exception des régions monocaténaires non codantes des télomères ). Le génome humain haploïde (23 chromosomes ) est estimé à environ 3,2 milliards de paires de bases et contient 20 000 à 25 000 gènes codant des protéines distincts. Un kilobase (kb) est une unité de mesure en biologie moléculaire égale à 1 000 paires de bases d'ADN ou d'ARN. Le nombre total de paires de bases d'ADN sur Terre est estimé à 5,0 de tonnes . En comparaison, la masse totale de la biosphère a été estimée à 4 TtC (milliards de tonnes de carbone ).
L’IUPAC ne formule aucune recommandation spécifique pour différencier les types de liaisons non covalentes. Lorsque cette distinction s’avère nécessaire, cet article utilise l’astérisque (*) pour désigner la paire de Hogsteen.
Liaisons hydrogène et stabilité
 |
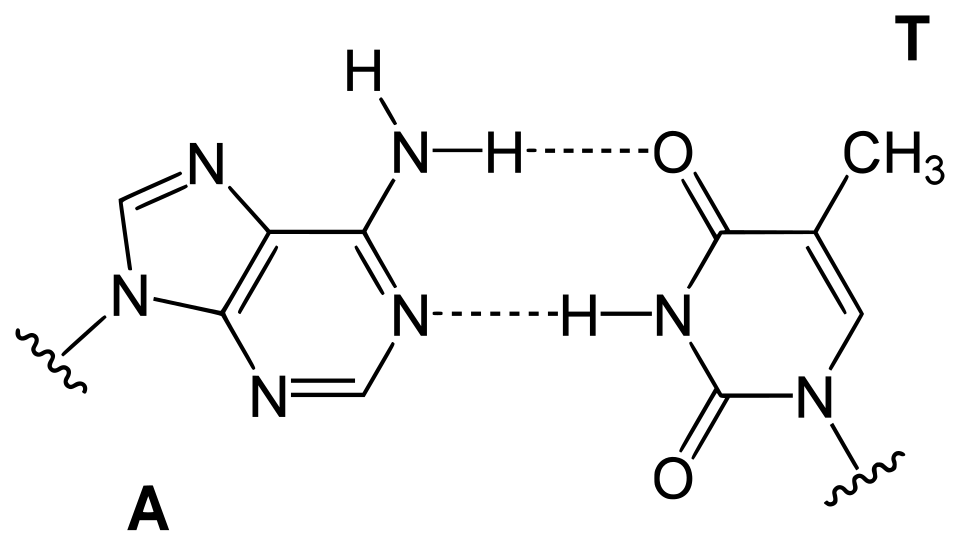 |
Les liaisons hydrogène constituent l'interaction chimique à la base des règles d'appariement des bases décrites précédemment. Une correspondance géométrique appropriée entre les donneurs et les accepteurs de liaisons hydrogène permet uniquement la formation stable des paires « correctes ». L'ADN à forte teneur en GC est plus stable que l'ADN à faible teneur en GC. Cependant, et c'est un point crucial, les interactions d'empilement sont principalement responsables de la stabilisation de la structure en double hélice ; la contribution de l'appariement des bases de Watson-Crick à la stabilité structurale globale est minime, mais son rôle dans la spécificité sous-jacente à la complémentarité est, en revanche, d'une importance capitale, car celle-ci sous-tend les processus dépendants de la matrice du dogme central (par exemple, la réplication de l'ADN ).
Les nucléobases les plus volumineuses , l'adénine et la guanine, appartiennent à la classe des purines, des structures chimiques à double cycle ; les nucléobases les plus petites, la cytosine et la thymine (ainsi que l'uracile), appartiennent à la classe des pyrimidines , des structures chimiques à simple cycle . Les purines sont complémentaires uniquement des pyrimidines : les appariements pyrimidine-pyrimidine sont énergétiquement défavorables car les molécules sont trop éloignées pour permettre la formation de liaisons hydrogène ; les appariements purine-purine sont également énergétiquement défavorables car les molécules sont trop proches, ce qui entraîne une répulsion par chevauchement. L'appariement des bases AT, GC ou UA (dans l'ARN) entre une purine et une pyrimidine conduit à une structure duplex correcte. Les seuls autres appariements purine-pyrimidine possibles sont AC, GT et UG (dans l'ARN) ; ces appariements sont incorrects car les motifs donneurs et accepteurs de liaisons hydrogène ne correspondent pas. L'appariement GU, avec deux liaisons hydrogène, se produit assez souvent dans l'ARN (voir paire de bases oscillantes ).
Les molécules d'ADN et d'ARN appariées sont relativement stables à température ambiante, mais les deux brins de nucléotides se séparent au-dessus d'un point de fusion déterminé par la longueur des molécules, le degré d'appariement incorrect (le cas échéant) et la teneur en GC. Une teneur en GC élevée entraîne une température de fusion plus élevée ; il n'est donc pas surprenant que les génomes d' organismes extrêmophiles tels que Thermus thermophilus soient particulièrement riches en GC. À l'inverse, les régions d'un génome qui doivent se séparer fréquemment — par exemple, les régions promotrices des gènes fréquemment transcrits — sont comparativement pauvres en GC (voir par exemple l'encadré TATA ). La teneur en GC et la température de fusion doivent également être prises en compte lors de la conception des amorces pour les réactions PCR .extrémité 5′ vers l' extrémité 3′ ; ainsi, le brin inférieur (brin complémentaire) est écrit de 3′ vers 5′.
- Une séquence d'ADN appariée en bases :
l'uracile est substitué à la thymine dans le brin d'ARN :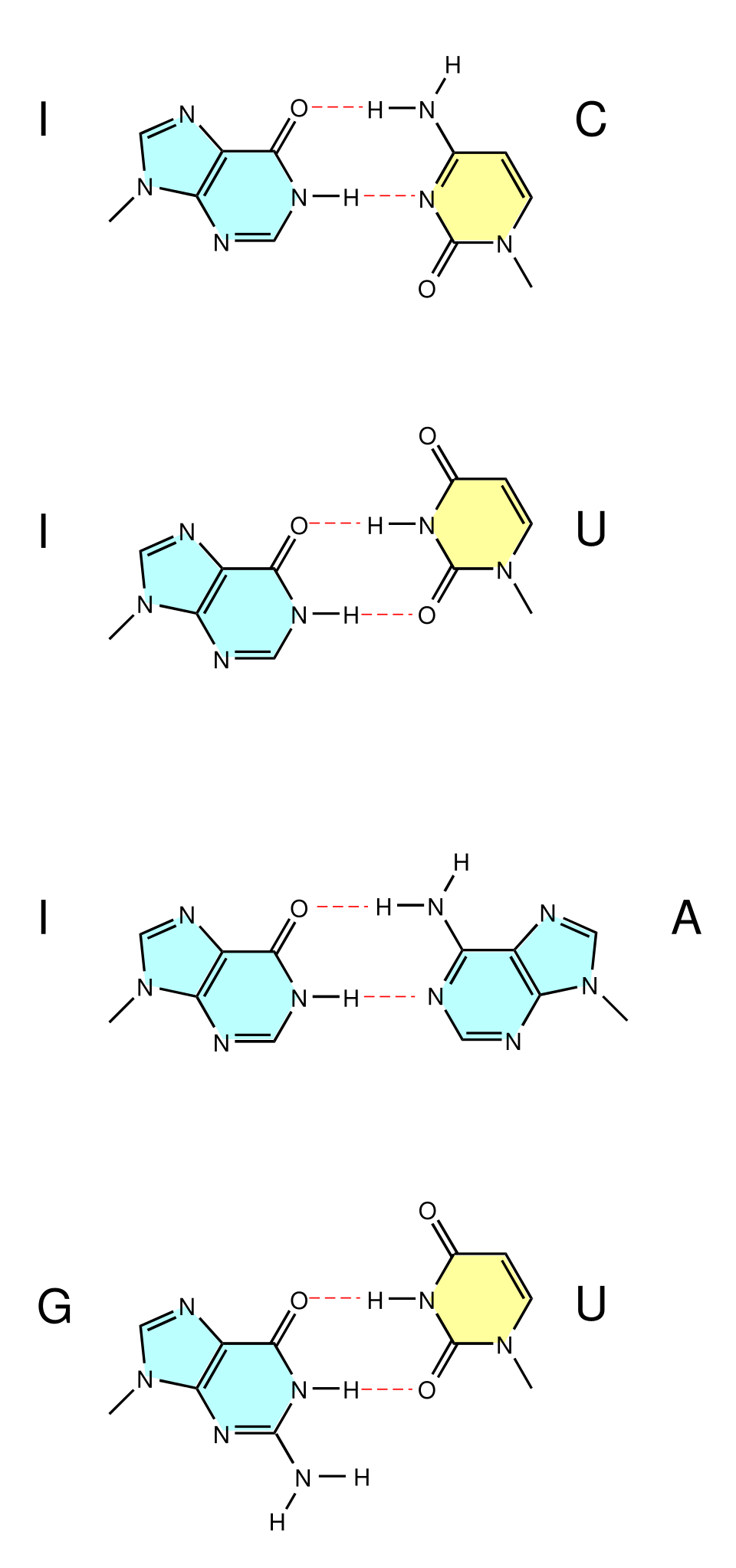 paires de bases oscillantes
paires de bases oscillantes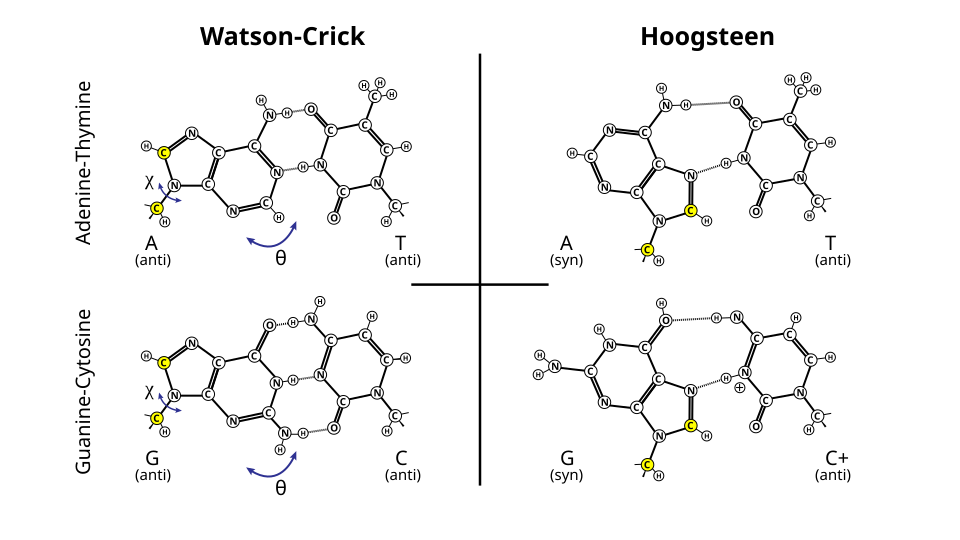 Comparaison des paires de bases de Hoogsteen et de Watson-Crick.
Comparaison des paires de bases de Hoogsteen et de Watson-Crick.
Outre l'appariement canonique de Watson-Crick (A•T/UG•C), certaines conditions peuvent également favoriser l'appariement de bases avec une orientation alternative, ainsi qu'un nombre et une géométrie différents des liaisons hydrogène. Ces appariements s'accompagnent de modifications de la conformation locale du squelette peptidique.appariement de bases oscillant qui se produit entre les ARNt et les ARNm à la troisième position de nombreux codons lors de la transcription et lors de la charge des ARNt par certaines ARNt synthétases . Ils ont également été observés dans les structures secondaires de certaines séquences d'ARN
De plus, l'appariement de bases de Hoogsteen (généralement noté A*U/T et G*C) peut se produire lorsqu'une face différente d'une base purique est utilisée pour l'appariement. Ce phénomène est observé dans certaines séquences d'ADN (par exemple, les dinucléotides CA et TA) en équilibre dynamique avec l'appariement Watson-Crick standard. Ces appariements ont également été observés dans certains complexes protéine-ADN. Il existe aussi un type d'appariement de bases de Hoogsteen inversé dans l'ARNt , où une purine et une pyrimidine utilisent chacune une face différente.
Outre ces appariements de bases alternatifs, une grande variété de liaisons hydrogène entre les bases est observée dans la structure secondaire et tertiaire de l'ARN. Ces liaisons sont souvent nécessaires à la forme précise et complexe d'un ARN, ainsi qu'à sa liaison à des partenaires d'interaction.
paires de bases et mutation
Réparation des incompatibilités
Des mésappariements de bases peuvent être générés par des erreurs de réplication de l'ADN et comme intermédiaires lors de la recombinaison homologue . Le processus de réparation des mésappariements doit généralement reconnaître et réparer correctement un petit nombre de mésappariements au sein d'une longue séquence d'ADN normale. Pour réparer les mésappariements formés lors de la réplication de l'ADN, plusieurs mécanismes de réparation spécifiques ont évolué afin de distinguer le brin matrice du brin néoformé et de n'éliminer que le nucléotide incorrect nouvellement inséré (afin d'éviter une mutation) . Les protéines impliquées dans la réparation des mésappariements lors de la réplication de l'ADN, ainsi que la signification clinique des anomalies de ce processus, sont décrites dans l'article « Réparation des mésappariements de l'ADN » . Le processus de correction des mésappariements lors de la recombinaison est décrit dans l'article « Conversion génique » .
analogues de base et intercalants
D’autres substances chimiques, appelées intercalants d’ADN , s’insèrent dans l’espace entre deux bases adjacentes sur un seul brin et induisent des mutations par décalage du cadre de lecture en se faisant passer pour une base, ce qui amène le mécanisme de réplication de l’ADN à sauter des bases ou à insérer des nucléotides supplémentaires au niveau du site d’intercalation. La plupart des intercalants sont de gros composés polyaromatiques et sont des cancérogènes avérés ou suspectés . On peut citer comme exemples le bromure d’éthidium et l’acridine .
En tant qu'unité de longueur
- bp = paire de bases — une bp correspond à environ 3,4 Å (340 pm ) de longueur le long du brin, et à environ 618 ou 643 daltons pour l'ADN et l'ARN respectivement.
- kb (= kbp) = kilo-paire de bases = 1 000 paires de bases
- Mb (= Mbp) = méga-paire de bases = 1 000 000 pb
- Gb (= Gbp) = giga-paire de bases = 1 000 000 000 paires de bases
Pour l'ADN/ARN simple brin, on utilise les nucléotides comme unités — abrégés nt (ou knt, Mnt, Gnt) — car ils ne sont pas appariés. Afin de distinguer les unités de stockage informatique des bases, on peut utiliser kbp, Mbp, Gbp, etc. pour les paires de bases.
Le centimorgan est également souvent utilisé pour indiquer une distance le long d'un chromosome, mais le nombre de paires de bases auquel il correspond varie considérablement selon les schémas de recombinaison chromosomique . Dans le génome humain, le centimorgan représente environ 1 million de paires de bases.
Paire de bases non naturelles (UBP)
En 2002, l'équipe d'Ichiro Hirao au Japon a développé une paire de bases non naturelle entre la 2-amino-8-(2-thiényl)purine (s) et la pyridine-2-one (y), fonctionnelle lors de la transcription et de la traduction, permettant l'incorporation site-spécifique d'acides aminés non conventionnels dans les protéines. En 2006, ils ont créé la 7-(2-thiényl)imidazo[4,5-b]pyridine (Ds) et le pyrrole-2-carbaldéhyde (Pa) comme troisième paire de bases pour la réplication et la transcription. Par la suite, Ds et le 4-[3-(6-aminohexanamido)-1-propynyl]-2-nitropyrrole (Px) ont été découverts comme une paire à haute fidélité pour l'amplification par PCR. En 2013, ils ont appliqué la paire Ds-Px à la génération d'aptamères d'ADN par sélection in vitro (SELEX) et ont démontré que l'expansion de l'alphabet génétique augmentait significativement les affinités des aptamères d'ADN pour les protéines cibles.
En 2012, une équipe de scientifiques américains dirigée par Floyd Romesberg, biologiste chimiste au Scripps Research Institute de San Diego, en Californie, a publié la conception d'une paire de bases non naturelle (PBN). Ces deux nouveaux nucléotides artificiels, ou paires de bases non naturelles (PBN), ont été nommés d5SICS et dNaM . Plus précisément, ces nucléotide artificiels, composés de nucléobases hydrophobes , présentent deux cycles aromatiques fusionnés formant un complexe (d5SICS–dNaM), ou paire de bases, dans l'ADN. L'équipe a conçu divers modèles in vitro, ou « tubes à essai », contenant cette paire de bases non naturelle et a confirmé sa réplication efficace et fidèle dans pratiquement tous les contextes de séquence, grâce aux techniques in vitro standard modernes , notamment l'amplification de l'ADN par PCR et les applications basées sur la PCR. Leurs résultats montrent que pour la PCR et les applications basées sur la PCR, la paire de bases non naturelles d5SICS–dNaM est fonctionnellement équivalente à une paire de bases naturelles, et que lorsqu'elle est combinée avec les deux autres paires de bases naturelles utilisées par tous les organismes, A–T et G–C, elle fournit un « alphabet génétique » à six lettres entièrement fonctionnel et étendu.
En 2014, la même équipe du Scripps Research Institute a rapporté avoir synthétisé un fragment d'ADN circulaire, appelé plasmide, contenant les paires de bases naturelles TA et CG, ainsi que la paire de bases non naturelles (UBP) la plus performante. Le laboratoire de Romesberg l'avait conçu et inséré dans des cellules de la bactérie Escherichia coli (E. coli) , qui a réussi à répliquer les paires de bases non naturelles sur plusieurs générations. La transfection n'a pas entravé la croissance des cellules d' E. coli et n'a montré aucun signe de perte des paires de bases non naturelles par les mécanismes naturels de réparation de l'ADN . Il s'agit du premier exemple connu d'un organisme vivant transmettant un code génétique étendu aux générations suivantes. Romesberg a indiqué que lui et ses collègues avaient créé 300 variantes pour affiner la conception des nucléotides afin qu'ils soient suffisamment stables et puissent être répliqués aussi facilement que les nucléotides naturels lors de la division cellulaire. Ce résultat a été obtenu en partie grâce à l'ajout d'un gène algal auxiliaire exprimant un transporteur de nucléotides triphosphates qui importe efficacement les triphosphates de d5SICSTP et de dNaMTP dans la bactérie E. coli . Les voies de réplication bactériennes naturelles les utilisent ensuite pour répliquer fidèlement un plasmide contenant d5SICS-dNaM. D'autres chercheurs ont été surpris de constater que les bactéries répliquaient ces sous-unités d'ADN synthétiques.
L'incorporation réussie d'une troisième paire de bases constitue une avancée majeure vers l'objectif d'accroître considérablement le nombre d' acides aminés pouvant être codés par l'ADN, passant des 20 acides aminés actuels à un nombre théoriquement possible de 172, augmentant ainsi le potentiel des organismes vivants à produire de nouvelles protéines . Ces séquences d'ADN artificielles ne codent encore pour rien, mais les scientifiques envisagent qu'elles pourraient être conçues pour fabriquer de nouvelles protéines ayant des applications industrielles ou pharmaceutiques. Selon des experts, l'ADN synthétique intégrant cette paire de bases non naturelle ouvre la voie à l'existence de formes de vie basées sur un code génétique différent.
Sources de données pour les forces des paires de bases
- Fragments d'hélice tels que AA•UU et GGUC•CUGG (la séquence à gauche du deux-points est dans le sens habituel 5' vers 3', mais celle à droite est écrite dans le sens inverse 3' vers 5')
- mésappariements terminaux (non-paires à l'extrémité des hélices, par exemple CA•GA).
Une liste de modèles de « plus proches voisins » peut être trouvée dans la
Revenez a l index pour explorer davantage de pages sur l histoire, la science, la culture, la geographie et la societe en francais.Plus d articles de Worldlex Wiki