Une mutation ponctuelle est une mutation génétique où une seule base nucléotidique est modifiée, insérée ou supprimée dans une séquence d'ADN ou d'ARN du génome d'un organisme. Les mutations ponctuelles ont divers effets sur la protéine produite ; ces conséquences sont relativement prévisibles en fonction des spécificités de la mutation. Elles peuvent être nulles (par exemple, les mutations synonymes ) ou délétères (par exemple, les mutations par décalage du cadre de lecture ), affectant la production, la composition et la fonction de la protéine.
la réplication de l'ADN . La réplication de l'ADN a lieu lorsqu'une molécule d'ADN double brin crée deux brins d'ADN simple brin, chacun servant de matrice pour la synthèse du brin complémentaire. Une seule mutation ponctuelle peut modifier la séquence d'ADN entière. Le remplacement d'une purine ou d'une pyrimidine peut entraîner la modification de l'acide aminé codé par les nucléotides .Les mutations ponctuelles peuvent résulter de mutations spontanées survenant lors de la réplication de l'ADN . Le taux de mutation peut être accru par des mutagènes . Ces mutagènes peuvent être physiques, comme les rayonnements UV , les rayons X ou la chaleur extrême, ou chimiques (molécules qui déplacent les paires de bases ou perturbent la structure en hélice de l'ADN). Les mutagènes associés aux cancers sont souvent étudiés afin de mieux comprendre le cancer et sa prévention.
Il existe plusieurs mécanismes à l'origine des mutations ponctuelles. Premièrement, les rayons ultraviolets (UV) et les rayonnements de haute fréquence ont un pouvoir ionisant, ce qui peut affecter l'ADN. Les molécules d'oxygène réactives, associées aux radicaux libres (sous-produits du métabolisme cellulaire), peuvent également endommager l'ADN. Ces réactifs peuvent provoquer des cassures simple brin et double brin. Troisièmement, la dégradation des liaisons de l'ADN constitue un autre obstacle au maintien de son intégrité. Enfin, des erreurs de réplication peuvent entraîner des mutations par substitution, insertion ou délétion.
catégorisation
catégorisation transition/transversion
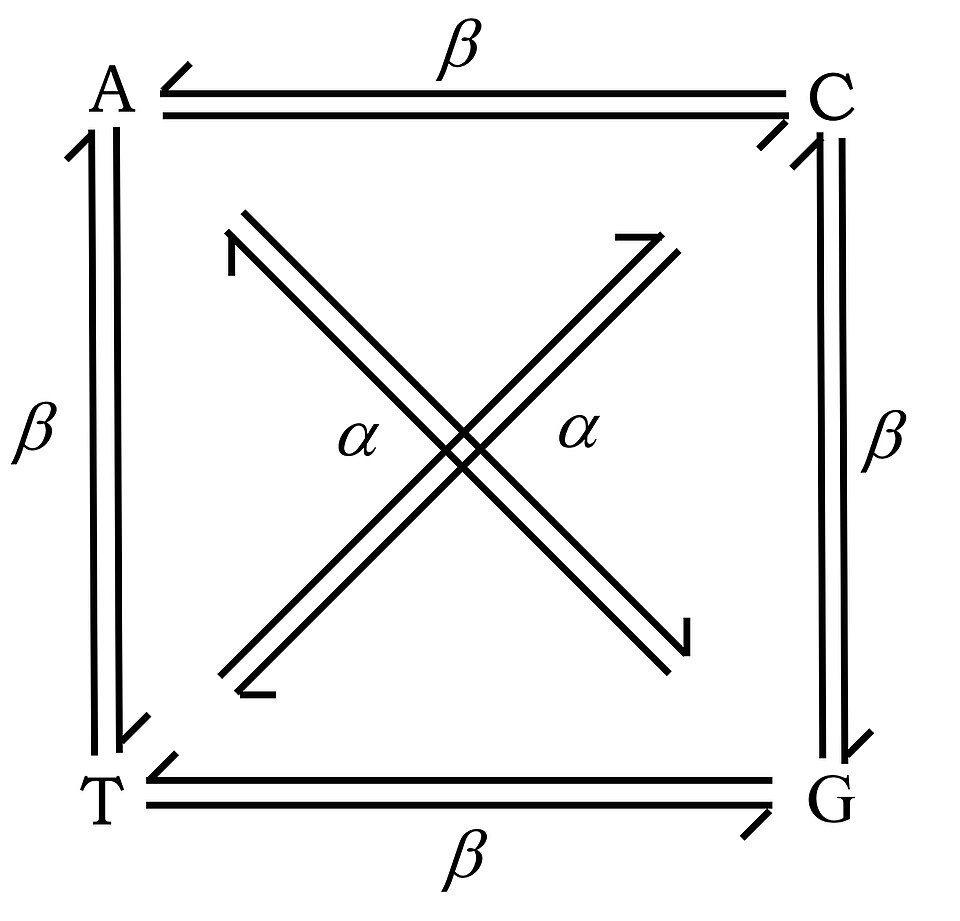
En 1959, Ernst Freese a introduit les termes « transitions » et « transversions » pour catégoriser différents types de mutations ponctuelles. Les transitions correspondent au remplacement d'une base purique par une autre purine ou d'une pyrimidine par une autre pyrimidine. Les transversions correspondent au remplacement d'une purine par une pyrimidine ou inversement. Il existe une différence systématique de fréquence entre les transitions (α) et les transversions (β). Les mutations par transition sont environ dix fois plus fréquentes que les transversions.
catégorisation fonctionnelle
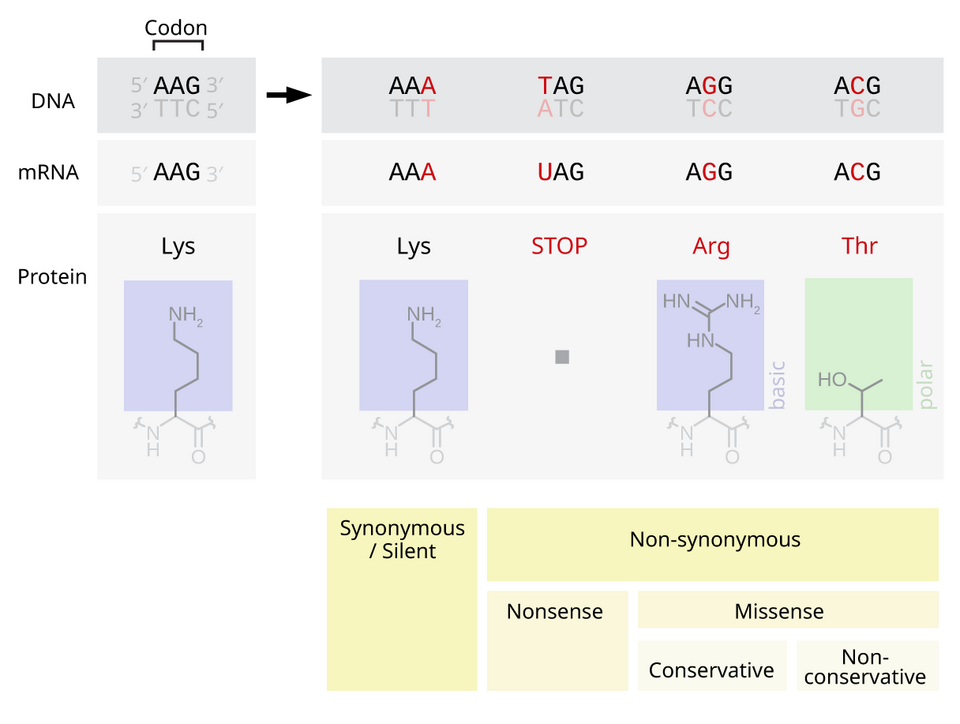
Les mutations non-sens comprennent les mutations par gain de codon stop et par perte de codon d'initiation. Une mutation par gain de codon stop entraîne l' apparition prématurée d'un codon stop ( un codon stop est ajouté ), signalant la fin de la traduction. Cette interruption provoque un raccourcissement anormal de la protéine. Le nombre d'acides aminés perdus détermine l'impact sur la fonctionnalité de la protéine, voire sa capacité à fonctionner. Une mutation par perte de codon stop est une mutation affectant le codon stop initial ( un codon stop est supprimé ), entraînant une extension anormale de l'extrémité C-terminale de la protéine. Une mutation par gain de codon d'initiation crée un codon d'initiation AUG en amont du site d'initiation initial. Si ce nouveau codon AUG est proche du site d'initiation initial, en phase dans le transcrit mature et en aval d'un site de liaison ribosomique, il peut être utilisé pour initier la traduction. L'effet probable est l'ajout d'acides aminés supplémentaires à l'extrémité N-terminale de la protéine. Des mutations par décalage du cadre de lecture sont également possibles lors d'une mutation par gain de codon d'initiation, mais n'affectent généralement pas la traduction de la protéine initiale. Une mutation par perte de codon d'initiation est une mutation ponctuelle affectant le codon d'initiation AUG d'un transcrit, entraînant une réduction ou une élimination de la production de la protéine.
Les mutations faux-sens codent pour un acide aminé différent. Une mutation faux-sens modifie un codon, ce qui entraîne la création d'une protéine différente ; il s'agit d'une mutation non synonyme. Les mutations conservatrices entraînent un changement d'acide aminé. Cependant, les propriétés de cet acide aminé restent inchangées (par exemple, hydrophobe, hydrophile, etc.). Parfois, la modification d'un seul acide aminé dans la protéine n'est pas préjudiciable à l'organisme dans son ensemble. La plupart des protéines peuvent supporter une ou deux mutations ponctuelles avant que leur fonction ne soit altérée. Les mutations non conservatrices entraînent un changement d'acide aminé dont les propriétés diffèrent de celles de la protéine sauvage . La protéine peut perdre sa fonction, ce qui peut provoquer une maladie chez l'organisme. Par exemple, la drépanocytose est causée par une mutation ponctuelle unique (une mutation faux-sens) dans le gène de la bêta- hémoglobine , qui convertit un codon GAG en GUG, codant pour la valine au lieu de l'acide glutamique . La protéine peut également présenter un « gain de fonction » ou être activée, comme c'est le cas lors de la mutation remplaçant une valine par un acide glutamique dans le gène BRAF ; ceci entraîne l'activation de la protéine RAF, provoquant une signalisation proliférative illimitée dans les cellules cancéreuses. Ce sont deux exemples de mutation non conservative (faux-sens).
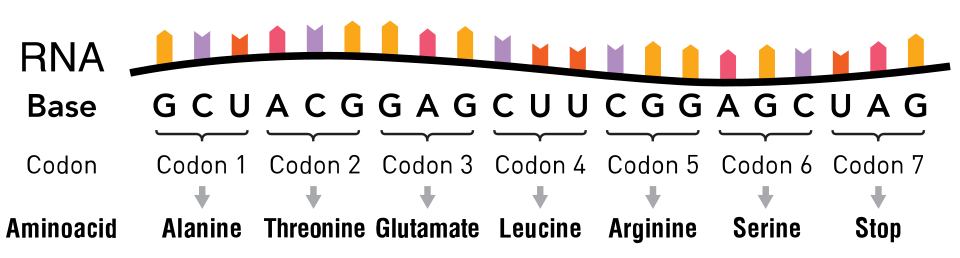
Les mutations silencieuses codent pour le même acide aminé ( substitution synonyme ). Une mutation silencieuse n'affecte pas la fonction de la protéine . Un seul nucléotide peut changer, mais le nouveau codon spécifie le même acide aminé, ce qui donne une protéine non mutée. Ce type de changement est appelé mutation synonyme car l'ancien et le nouveau codon codent pour le même acide aminé. Ceci est possible car 64 codons ne spécifient que 20 acides aminés. Des codons différents peuvent cependant entraîner des niveaux d'expression protéique différents.
Insertions et délétions d'une seule paire de bases
Parfois, le terme mutation ponctuelle est utilisé pour décrire les insertions ou les délétions d'une seule paire de bases (qui ont un effet plus néfaste sur la protéine synthétisée car les nucléotides sont toujours lus en triplets, mais dans des cadres différents : une mutation appelée mutation par décalage du cadre de lecture ).
Conséquences générales
Les mutations ponctuelles survenant dans des séquences non codantes sont le plus souvent sans conséquence, bien qu'il existe des exceptions. Si la paire de bases mutée se situe dans la séquence promotrice d'un gène, l'expression de ce gène peut être modifiée. De même, si la mutation se produit au niveau du site d'épissage d'un intron , elle peut perturber l'épissage correct de l' ARN pré-messager transcrit .
La modification d'un seul acide aminé peut entraîner le changement de l' ensemble du peptide et, par conséquent, de la protéine entière. Cette nouvelle protéine est appelée germinales peuvent engendrer des caractéristiques bénéfiques ou néfastes, voire des maladies. Elles induisent des adaptations en fonction de l'environnement. Une mutation avantageuse confère un avantage à l'organisme et permet la transmission de ce caractère de génération en génération, améliorant ainsi la population. La théorie scientifique de l'évolution repose en grande partie sur les mutations ponctuelles cellulaires . Cette théorie explique la diversité et l'histoire du vivant sur Terre. Elle stipule que les mutations bénéfiques permettent à l'organisme de prospérer et de se reproduire, transmettant ainsi ses gènes mutés à la génération suivante. À l'inverse, les mutations néfastes entraînent la mort de l'organisme ou réduisent ses chances de reproduction, un phénomène connu sous le nom de sélection naturelle .
Les mutations peuvent avoir différents effets à court et à long terme. Parmi les effets mineurs, on peut citer l'arrêt du cycle cellulaire à plusieurs étapes. Cela signifie qu'un codon codant pour l'acide aminé glycine peut être transformé en codon stop, ce qui entraîne la production de protéines déformées et incapables de remplir leur fonction. Comme les mutations peuvent affecter l'ADN et donc la chromatine , elles peuvent empêcher la mitose en raison de l'absence d'un chromosome complet. Des problèmes peuvent également survenir lors de la transcription et de la réplication de l'ADN. Tous ces facteurs empêchent la reproduction cellulaire et conduisent ainsi à la mort de la cellule. Les effets à long terme peuvent inclure une modification permanente d'un chromosome, qui peut être à l'origine d'une mutation. Ces mutations peuvent être bénéfiques ou délétères. Le cancer est un exemple de mutation délétère.
D'autres effets des mutations ponctuelles, ou polymorphismes nucléotidiques simples (SNP) de l'ADN, dépendent de leur localisation au sein du gène. Par exemple, si la mutation survient dans la région codante du gène, la séquence d'acides aminés de la protéine codée peut être altérée, entraînant une modification de sa fonction, de sa localisation, de sa stabilité ou de celle du complexe protéique. De nombreuses méthodes ont été proposées pour prédire les effets des mutations faux-sens sur les protéines. Les algorithmes d'apprentissage automatique entraînent leurs modèles à distinguer les mutations connues associées à des maladies des mutations neutres, tandis que d'autres méthodes n'entraînent pas explicitement leurs modèles. Cependant, la quasi-totalité des méthodes exploitent la conservation évolutive, en supposant que les modifications aux positions conservées tendent à être plus délétères. Si la majorité des méthodes proposent une classification binaire des effets des mutations en effets délétères et bénins, un niveau d'annotation plus poussé est nécessaire pour expliquer pourquoi et comment ces mutations endommagent les protéines.
De plus, si la mutation survient dans la région du gène où le complexe de transcription se lie à la protéine, elle peut affecter la liaison des facteurs de transcription, car les courtes séquences nucléotidiques reconnues par ces facteurs seront altérées. Les mutations dans cette région peuvent affecter l'efficacité de la transcription génique, ce qui peut modifier les niveaux d'ARNm et, par conséquent, les niveaux de protéines en général.
Les mutations ponctuelles peuvent avoir plusieurs effets sur le comportement et la reproduction d'une protéine, selon leur position dans la séquence d'acides aminés. Si la mutation survient dans la région du gène codant pour la protéine, l'acide aminé correspondant peut être modifié. Ce léger changement dans la séquence d'acides aminés peut entraîner une modification de la fonction, de l'activation de la protéine (c'est-à-dire sa capacité à se lier à une enzyme donnée), de sa localisation intracellulaire ou encore de la quantité d'énergie libre qu'elle contient.
Si la mutation survient dans la région du gène où le complexe de transcription se lie à la protéine, elle peut affecter la façon dont les facteurs de transcription se lient à cette protéine. Les mécanismes de transcription se lient à une protéine par reconnaissance de courtes séquences nucléotidiques. Une mutation dans cette région peut altérer ces séquences et, par conséquent, modifier la façon dont les facteurs de transcription se lient à la protéine. Les mutations dans cette région peuvent affecter l'efficacité de la transcription génique, qui contrôle à la fois les niveaux d'ARNm et les niveaux protéiques totaux.
Maladies spécifiques causées par des mutations ponctuelles
Les mutations ponctuelles — modifications d'une seule base dans la séquence d'ADN — constituent l'une des causes moléculaires les plus fréquentes de maladies humaines. En altérant un seul nucléotide, ces mutations peuvent substituer un acide aminé par un autre, introduire des codons stop prématurés ou perturber les signaux d'épissage normaux. Selon leur localisation et leur impact sur la protéine codée, les mutations ponctuelles peuvent abolir l'activité enzymatique, déstabiliser des domaines structuraux ou perturber les interactions régulatrices. Dans de nombreuses maladies héréditaires, une simple substitution faux-sens ou non-sens suffit à déclencher une cascade de défaillances biochimiques, entraînant des symptômes précoces ou persistants. Dans le cancer, les mutations ponctuelles somatiques peuvent inactiver les gènes suppresseurs de tumeurs ou hyperactiver les oncogènes, favorisant ainsi une croissance cellulaire incontrôlée.
Dans le paysage génétique humain, des milliers de maladies dues à des mutations ponctuelles ont été répertoriées, allant de troubles relativement fréquents comme la drépanocytose et la mucoviscidose à des syndromes extrêmement rares qui n'affectent qu'une poignée de familles dans le monde. Bien que chaque maladie présente ses propres particularités physiopathologiques, elles partagent un point commun : une modification localisée et précise de la séquence génique peut compromettre la fonction protéique d'une manière qu'aucun réarrangement chromosomique plus important ni aucune altération du nombre de copies ne pourrait impliquer. Les mutations ponctuelles étant souvent accessibles à des tests génétiques ciblés, elles soulignent également combien le diagnostic moléculaire et les thérapies personnalisées (par exemple, les petites molécules qui stabilisent une enzyme mutante) dépendent de la connaissance précise du codon altéré. Bien que les exemples suivants illustrent la diversité des maladies liées à des mutations ponctuelles, on en compte plus de 300 000 dans HGMD et plus d'un million dans ClinVar, reflétant le vaste spectre des mutations ponctuelles humaines.
Cancer
Des mutations ponctuelles dans plusieurs protéines suppresseurs de tumeurs provoquent le cancer . Par exemple, des mutations ponctuelles dans le gène de la polypose adénomateuse familiale (APC) favorisent la tumorigenèse. Un nouveau test, la protéolyse parallèle rapide (FASTpp) , pourrait permettre un dépistage rapide des défauts de stabilité spécifiques chez les patients atteints de cancer.
neurofibromatose
Anémie falciforme
| août | GUG | CAC | CUG | ACU | USC | G A G | GAG | AAG | UCU | CCG | GUU | ACU |
| COMMENCER | Val | Son | Leu | Thr | Pro | Glu | Glu | Lys | Ser | Ala | Val | Thr |
| août | GUG | CAC | CUG | ACU | USC | G U G | GAG | AAG | UCU | CCG | GUU | ACU |
| COMMENCER | Val | Son | Leu | Thr | Pro | Val | Glu | Lys | Ser | Ala | Val | Thr |
maladie de Tay-Sachs
La maladie de Tay-Sachs est due à une anomalie génétique transmise des parents aux enfants. Cette anomalie se situe dans le gène HEXA, qui se trouve sur le chromosome 15.
Le gène HEXA code pour une partie d'une enzyme appelée bêta-hexosaminidase A, qui joue un rôle crucial dans le système nerveux. Cette enzyme contribue à la dégradation d'une substance grasse appelée ganglioside GM2 dans les cellules nerveuses. Les mutations du gène HEXA perturbent l'activité de la bêta-hexosaminidase A, empêchant ainsi la dégradation des substances grasses. Par conséquent, ces substances s'accumulent jusqu'à atteindre des niveaux mortels dans le cerveau et la moelle épinière. L'accumulation de ganglioside GM2 provoque des lésions progressives des cellules nerveuses. C'est l'origine des signes et symptômes de la maladie de Tay-Sachs.
mutation ponctuelle induite par la répétition
En biologie moléculaire , la mutation ponctuelle induite par répétition ( RIP) est un processus par lequel l'ADN accumule des mutations de transition G : C vers A : T . Des données génomiques indiquent que la RIP se produit ou s'est produite chez divers champignons , tandis que des données expérimentales montrent qu'elle est active chez *Neurospora crassa * , * Podospora anserina * , * Magnaporthe grisea * , * Leptosphaeria maculans * , *Gibberella zeae * , * Nectria haematococca et *Paecilomyces variotii . Chez *Neurospora crassa* , les séquences mutées par RIP sont souvent méthylées de novo .
La RIP (reconnaissance des séquences répétées) survient lors de la phase sexuée dans les noyaux haploïdes après la fécondation mais avant la réplication méiotique de l'ADN . Chez Neurospora crassa , les séquences répétées d'au moins 400 paires de bases sont sensibles à la RIP. Des répétitions présentant une identité nucléotidique aussi faible que 80 % peuvent également y être sujettes. Bien que le mécanisme exact de reconnaissance et de mutagénèse des répétitions soit encore mal compris, la RIP induit de multiples mutations de transition dans les séquences répétées .
Les mutations RIP ne semblent pas se limiter aux séquences répétées. En effet, par exemple, chez le champignon phytopathogène L. maculans , des mutations RIP sont observées dans des régions à copie unique, adjacentes aux éléments répétés. Ces régions sont soit des régions non codantes, soit des gènes codant pour de petites protéines sécrétées, notamment des gènes d'avirulence. Le degré de RIP au sein de ces régions à copie unique était proportionnel à leur proximité avec les éléments répétitifs.
Rep et Kistler ont émis l'hypothèse que la présence de régions hautement répétitives contenant des transposons pourrait favoriser la mutation des gènes effecteurs résidents. Ainsi, la présence de gènes effecteurs au sein de telles régions favoriserait leur adaptation et leur diversification lorsqu'ils sont exposés à une forte pression de sélection.
Comme la mutation RIP est généralement observée comme étant restreinte aux régions répétitives et non aux régions à copie unique, Fudal et al. ont suggéré qu'une diffusion de la mutation RIP pourrait se produire à une distance relativement courte d'une répétition affectée par RIP. Ce phénomène a effectivement été rapporté chez N. crassa, où une diffusion de RIP a été détectée dans des séquences à copie unique situées à au moins 930 pb de la limite de séquences dupliquées voisines . L'élucidation du mécanisme de détection des séquences répétées conduisant à la mutation RIP pourrait permettre de comprendre comment les séquences flanquantes peuvent également être affectées.
Mécanisme
Le système RIP induit des mutations de transition G : C vers A : T au sein des séquences répétées. Cependant, le mécanisme de détection de ces séquences reste inconnu. RID est la seule protéine connue essentielle à RIP. Il s'agit d'une protéine de type ADN méthyltransférase dont la mutation ou l'inactivation entraîne la perte de RIP . La délétion de l' homologue de rid chez Aspergillus nidulans , dmtA , provoque une perte de fertilité , tandis que la délétion de l' homologue de rid chez méthylation induite pré-méiotiquement (MIP) .
Conséquences
On pense que le mécanisme RIP a évolué comme mécanisme de défense contre les éléments transposables , qui s'apparentent à des parasites en envahissant et en se multipliant au sein du génome. Le mécanisme RIP génère de multiples mutations faux-sens et non-sens dans la séquence codante. Cette hypermutation de GC en AT dans les séquences répétitives élimine les produits géniques fonctionnels de la séquence (s'il y en avait initialement). De plus, de nombreux nucléotides contenant de la cytosine sont méthylés , ce qui diminue la transcription.
Utilisation en biologie moléculaire
L’efficacité du système RIP pour détecter et muter les séquences répétées a été démontrée par les biologistes travaillant sur Neurospora crassa , qui l’ont utilisé comme outil de mutagenèse . Une seconde copie d’un gène initialement unique est d’abord introduite dans le génome . Le champignon doit ensuite se reproduire et achever son cycle sexuel pour activer le système RIP. Une seule fécondation permet d’obtenir de nombreuses mutations différentes au sein du gène dupliqué, ce qui conduit à l’obtention d’allèles inactivés, généralement par des mutations non-sens , ainsi que d’allèles porteurs de mutations faux-sens .
Histoire
Hertwig étudia les oursins et remarqua que chaque ovule contenait un noyau avant la fécondation et deux après. Cette découverte prouva qu'un seul spermatozoïde pouvait féconder un ovule et confirma ainsi le processus de méiose. Hermann Fol poursuivit les recherches de Hertwig en testant les effets de l'injection de plusieurs spermatozoïdes dans un ovule et constata que le processus ne fonctionnait pas avec plus d'un spermatozoïde.
Flemming a commencé ses recherches sur la division cellulaire en 1868. L'étude des cellules était un sujet de plus en plus populaire à cette époque. Dès 1873, Schneider avait commencé à décrire les étapes de la division cellulaire. Flemming a approfondi cette description en 1874 et 1875 en expliquant ces étapes plus en détail. Il a également contesté les conclusions de Schneider selon lesquelles le noyau se divisait en structures en forme de bâtonnets, en suggérant que le noyau se divisait en réalité en filaments qui se séparaient ensuite. Flemming a conclu que les cellules se répliquent par division cellulaire, plus précisément par mitose.
Matthew Meselson et Franklin Stahl sont reconnus pour avoir découvert la réplication de l'ADN . Watson et Crick avaient déjà constaté que la structure de l'ADN suggérait l'existence d'un processus de réplication. Cependant, peu de recherches ont été menées sur cet aspect de l'ADN avant leurs travaux. Toutes les méthodes possibles pour déterminer le mécanisme de réplication de l'ADN ont été envisagées, mais aucune n'a abouti avant les recherches de Meselson et Stahl. Ces derniers ont introduit un isotope lourd dans de l'ADN et ont suivi sa distribution. Grâce à cette expérience, Meselson et Stahl ont pu démontrer que l'ADN se reproduit de manière semi-conservative.