L'éclatement , ou déclenchement en rafale , est un phénomène général extrêmement diversifié des schémas d'activation des neurones du système nerveux central et de la moelle épinière où des périodes de pics de potentiel d'action rapide sont suivies de périodes de repos beaucoup plus longues que les intervalles inter-pics typiques. On pense que l'éclatement est important dans le fonctionnement de générateurs de schémas centraux robustes [ la transmission de codes neuronaux [ et certaines neuropathologies telles que l'épilepsie . L'étude de l'éclatement, à la fois directement et dans la façon dont il participe à d'autres phénomènes neuronaux, est très populaire depuis les débuts des neurosciences cellulaires et est étroitement liée aux domaines de la synchronisation neuronale , du codage neuronal , de la plasticité et de l'attention .
Les rafales observées sont nommées en fonction du nombre de potentiels d'action discrets qui les composent : un doublet est une rafale à deux pics, un triplet en a trois et un quadruplet en a quatre. Les neurones qui sont intrinsèquement enclins à un comportement de rafale sont appelés « éclateurs » et cette tendance à éclater peut être le produit de l'environnement ou du phénotype de la cellule.
Contexte physiologique
Aperçu
Les neurones fonctionnent généralement en déclenchant des pics de potentiel d'action uniques de manière relativement isolée, tandis que des potentiels postsynaptiques d'entrée discrets se combinent et poussent le potentiel de membrane à travers le seuil . L'éclatement peut au contraire se produire pour de nombreuses raisons, mais les neurones peuvent généralement être regroupés comme présentant un éclatement induit par l'entrée ou un éclatement intrinsèque . La plupart des cellules présenteront un éclatement si elles sont entraînées par une entrée constante, sous le seuil et certaines cellules qui sont génotypiquement sujettes à l'éclatement (appelées éclateurs ) ont des systèmes de rétroaction complexes qui produiront des schémas d'éclatement avec moins de dépendance à l'entrée et parfois même de manière isolée.
Dans chaque cas, le système physiologique est souvent action de deux sous-systèmes liés. Le sous-système rapide est responsable de chaque pic produit par le neurone. Le sous-système lent module la forme et l'intensité de ces pics avant de finalement déclencher la quiescence.
L'éclatement piloté par l'entrée encode souvent l'intensité de l'entrée dans la fréquence d'éclatement où un neurone agit alors comme un intégrateur . L'éclatement intrinsèque est un phénomène plus spécialisé et est considéré comme jouant un rôle beaucoup plus diversifié dans le calcul neuronal .
Sous-système rapide
Sous-système lent
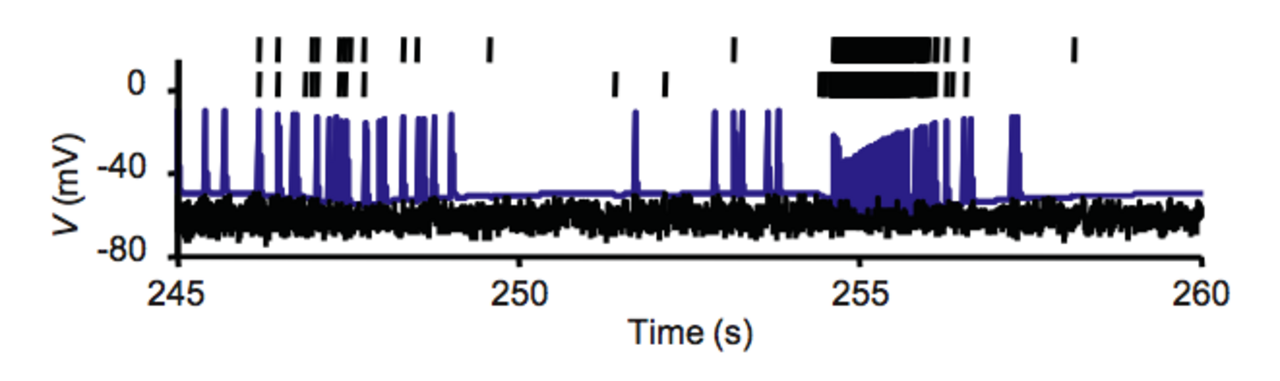
Les salves diffèrent des décharges toniques , généralement associées à des temps de pointe distribués selon la loi de Poisson pour un taux de décharge moyen donné, dans la mesure où les salves impliquent un « sous-système lent » physiologique qui finit par s'épuiser à mesure que les salves se poursuivent et doit ensuite être réapprovisionné avant que la cellule ne puisse à nouveau éclater (cf. période réfractaire ). Pendant l'événement de salves, ce sous-système lent module le moment et l'intensité des pointes émises et est considéré comme important dans les aspects informatiques du modèle de salves résultant. Il existe de nombreux mécanismes découverts de sous-systèmes lents, notamment les courants voltage- et Ca 2+ -dépendants et l'interaction de pointes entre les dendrites et le corps cellulaire .
Le sous-système lent est également connecté aux schémas endogènes d'éclatement dans les neurones, où le schéma peut être maintenu complètement par un mécanisme interne sans aucune entrée synaptique . Ce processus repose également sur les canaux calciques , qui dépolarisent le neurone en permettant un afflux d' ions calcium . Tant que les concentrations internes d'ions calcium restent à un niveau élevé, le neurone continuera à subir des périodes de pics rapides. Cependant, des niveaux élevés d'ions calcium déclenchent également une cascade de seconds messagers dans la cellule qui diminue l'afflux de calcium et favorise l'efflux et la mise en tampon du calcium . Lorsque les concentrations de calcium diminuent, la période d'éclatement rapide cesse et la phase de quiescence commence. Lorsque les niveaux de calcium sont bas, les canaux calciques d'origine se rouvrent, redémarrant le processus et créant un schéma d'éclatement.
Détection statistique
De manière isolée ou dans des modèles mathématiques, les explosions peuvent être reconnues car l'environnement et l'état du neurone peuvent être soigneusement observés et modulés. Cependant, lors de l'observation de neurones dans la nature, les explosions peuvent être difficiles à distinguer des schémas de déclenchement normaux. Afin de reconnaître les schémas d'explosion dans ces contextes, des méthodes statistiques sont utilisées pour déterminer les paramètres de seuil.
Le phénomène de rafale est caractérisé par un coefficient de variation (CV) des intervalles entre pics (ISI) supérieur à un, ou par un facteur Fano du nombre de pics supérieur à un, car le phénomène de rafale conduit à des schémas de pics plus irréguliers qu'un processus de Poisson (qui a un CV et un facteur Fano égaux à un). Alternativement, le coefficient de corrélation sérielle de la séquence ISI est positif pour les schémas de rafale, car dans ce cas, les ISI courts ont tendance à être suivis par des ISI plus courts (au moins si les rafales se composent de plus de deux pics).
Modèles mathématiques
Le comportement des neurones est souvent modélisé comme des systèmes dynamiques non linéaires à compartiment unique , où les états des neurones représentent des quantités physiologiques telles que la tension membranaire, le flux de courant et les concentrations de divers ions intra et extracellulaires. Ces modèles prennent le plus souvent la forme singulièrement perturbée
- sous-système rapide :
- sous-système lent :
où et sont tous deux des relations de style Hodgkin–Huxley , est un vecteur représentant les paramètres de cellule pertinents pour le sous-système rapide, est un vecteur représentant les paramètres du sous-système de modulation lente et est le rapport des échelles de temps entre les sous-systèmes rapide et lent.
Les modèles de dynamique neuronale présentent généralement un certain nombre d' attracteurs stables et instables dans l'espace des phases qui représentent des états de repos. Lorsque le système est suffisamment perturbé par des stimuli d'entrée, il peut suivre un chemin de retour complexe vers l' attracteur stable représentant un potentiel d'action. Dans les neurones en mode rafale, ces espaces dynamiques bifurquent entre les modes de repos et d'éclatement en fonction de la dynamique du système lent. Ces deux bifurcations peuvent prendre de nombreuses formes et le choix de la bifurcation, du mode de repos à l'éclatement et de l'éclatement au mode de repos, peut affecter les aspects comportementaux de l'attracteur.
La classification complète des bifurcations quiescentes vers éclatement et éclatement vers quiescentes conduit à 16 formes courantes et 120 formes possibles si la dimensionnalité du sous-système rapide n'est pas contrainte. Parmi les 16 formes les plus courantes, quelques-unes sont bien étudiées.
Burster à ondes carrées
Le burster à onde carrée/homocline , également appelé burster à onde carrée, est ainsi nommé parce que la forme de la trace de tension pendant un burst ressemble à une onde carrée en raison des transitions rapides entre l'attracteur à l'état de repos et le cycle limite de pointe.
Objectifs
Le phénomène de rafale est un phénomène très général qui s'observe dans de nombreux contextes et dans de nombreux systèmes neuronaux. Pour cette raison, il est difficile de lui trouver une signification ou un but spécifique. En effet, il joue de nombreux rôles. Dans un circuit donné, les rafales observées peuvent jouer un rôle dans l'un ou l'ensemble des mécanismes suivants et peuvent avoir un impact encore plus sophistiqué sur le réseau.
Plasticité synaptique
Les forces synaptiques entre les neurones suivent des changements qui dépendent du timing des pics et de leur rafale. Pour les synapses excitatrices du cortex, l'association d'un potentiel d'action dans le neurone présynaptique avec une rafale dans le neurone postsynaptique conduit à une potentialisation à long terme de la force synaptique, tandis que l'association d'un potentiel d'action dans le neurone présynaptique avec une seule rafale dans le neurone postsynaptique conduit à une dépression à long terme de la force synaptique. Une telle dépendance de la plasticité synaptique sur les schémas de timing des pics est appelée plasticité dépendante des rafales. La plasticité dépendante des rafales est observée avec des variations dans plusieurs zones du cerveau.
Multiplexage et routage
Certains neurones, parfois appelés résonateurs , présentent une sensibilité à des fréquences d'entrée spécifiques et se déclenchent soit plus rapidement, soit exclusivement lorsqu'ils sont stimulés à cette fréquence. Les neurones intrinsèquement en rafale peuvent utiliser cet effet de filtrage passe-bande afin de coder des neurones de destination spécifiques et de multiplexer les signaux le long d'un seul axone . Plus généralement, en raison de la dépression et de la facilitation synaptiques à court terme , des synapses spécifiques peuvent être résonantes pour certaines fréquences et ainsi devenir des cibles spécifiques viables pour les cellules en rafale. Lorsqu'il est combiné à une plasticité à long terme dépendante des rafales, un tel multiplexage peut permettre aux neurones de coordonner la plasticité synaptique à travers des réseaux hiérarchiques.
Synchronisation
La synchronisation des rafales fait référence à l'alignement des périodes d'éclatement et de repos dans les neurones interconnectés. En général, si un réseau de neurones en rafale est lié, il finira par se synchroniser pour la plupart des types d'éclatement. La synchronisation peut également apparaître dans des circuits ne contenant aucun neurone intrinsèquement en rafale, mais son apparence et sa stabilité peuvent souvent être améliorées en incluant des cellules intrinsèquement en rafale dans le réseau. Étant donné que la synchronisation est liée à la plasticité et à la mémoire via la plasticité hebbienne et la potentialisation à long terme, l'interaction avec la plasticité et l'éclatement intrinsèque est très importante .
Contenu de l'information et robustesse du canal
En raison de la nature tout ou rien des potentiels d'action, les pics simples ne peuvent encoder des informations que dans leurs intervalles interpics (ISI). Il s'agit d'une méthode de transfert d'informations intrinsèquement peu fidèle car elle dépend d'un timing très précis et est sensible à la perte de signal bruyante : si un seul pic est mal synchronisé ou mal reçu au niveau de la synapse, cela conduit à une perte de codage potentiellement irrécupérable . Étant donné que les salves intrinsèques sont censées être dérivées par un mécanisme de calcul dans le sous-système lent, chacune peut représenter une quantité beaucoup plus importante d'informations sous la forme spécifique d'une seule salves, ce qui conduit à une transmission beaucoup plus robuste. Les modèles physiologiques montrent que pour une entrée donnée, les timings interpics et intersalves sont beaucoup plus variables que le timing de la forme de la salves elle-même ce qui implique également que le timing entre les événements est une manière moins robuste d'encoder des informations.
L'alphabet élargi pour la communication rendu possible par la prise en compte des modèles de rafales comme des signaux discrets permet une plus grande capacité de canal dans les communications neuronales et fournit une connexion populaire entre le codage neuronal et la théorie de l'information .
Exemple de circuits neuronaux éclatés
Hippocampe
Le subiculum , un composant de la formation hippocampique , est censé relayer les signaux provenant de l'hippocampe vers de nombreuses autres parties du cerveau. Afin d'exécuter cette fonction, il utilise des neurones intrinsèquement éclatants pour convertir des stimuli simples prometteurs en modèles d'éclats plus durables afin de mieux concentrer l'attention sur de nouveaux stimuli et d'activer des circuits de traitement importants. Une fois ces circuits activés, le signal subiculaire revient à un mode de pointe unique.
complexe pré-Bötzinger
Le complexe pré-Bötzinger (preBötC) est situé dans la moelle ventrolatérale et est censé générer le rythme sous-jacent aux efforts inspiratoires chez les mammifères. Étant donné que la fréquence à laquelle les poumons doivent fonctionner peut varier en fonction de la demande métabolique, l'activité du preBötC est modulée sur une large gamme de fréquences et est capable d'entraîner le système respiratoire à répondre à la demande métabolique. Bien que les neurones stimulateurs cardiaques ne nécessitent pas nécessairement de neurones à rafales intrinsèques le preBötC contient une population hétérogène de neurones à impulsions régulières et de neurones à rafales intrinsèques. On pense que les neurones à rafales intrinsèques rendent les oscillations du preBötC plus résistantes aux changements de fréquence et à la régularité des efforts inspiratoires
Cortex cérébelleux
Il a été proposé que les neurones de Purkinje cérébelleux aient deux modes d'éclatement distincts : entraînés de manière dendritique , par le Ca dendritique2+
pointes , et somatiquement entraînées, où le Na persistant+
le courant est l'initiateur de l'éclatement et le SK K+
le courant est le terminateur de l'éclatement. Les neurones de Purkinje peuvent utiliser ces formes d'éclatement dans le codage des informations vers les noyaux cérébelleux profonds .