La mémoire est la faculté de l' esprit par laquelle les données ou les informations sont encodées , stockées et récupérées en cas de besoin. Il s'agit de la conservation d'informations au fil du temps afin d'influencer les actions futures . Si les événements passés ne pouvaient être remémorés, le langage, les relations et l'identité personnelle seraient impossibles à développer. La perte de mémoire est généralement décrite comme un oubli ou un trouble tel que l'amnésie .
La mémoire est souvent perçue comme un système de traitement de l'information, doté d'un fonctionnement explicite et implicite, composé d'un processeur sensoriel , d'une mémoire à court terme (ou mémoire de travail ) et d'une mémoire à long terme . Ce système peut être comparé au neurone . Le processeur sensoriel permet de percevoir les informations provenant du monde extérieur sous forme de stimuli chimiques et physiques et d'y porter une attention plus ou moins soutenue. La mémoire de travail assure l'encodage et la récupération des informations. Les informations, sous forme de stimuli, sont encodées par la mémoire de travail selon des fonctions explicites ou implicites. La mémoire de travail permet également de récupérer les informations stockées précédemment. Enfin, la mémoire à long terme a pour fonction de stocker les informations à travers différents modèles ou systèmes catégoriels
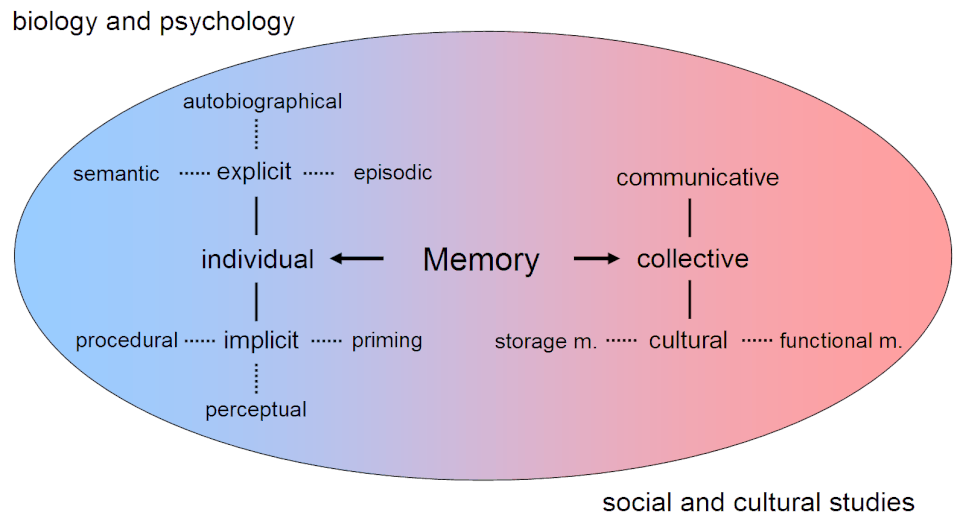
La mémoire déclarative, ou explicite , correspond au stockage et à la remémoration conscients des données. Elle englobe la mémoire sémantique et la mémoire épisodique . La mémoire sémantique désigne les informations encodées avec une signification spécifique. La mémoire épisodique, quant à elle, renvoie aux informations encodées selon un plan spatio-temporel. La mémoire déclarative est généralement le premier processus auquel on pense lorsqu'on évoque la mémoire. La mémoire non déclarative, ou implicite, correspond au stockage et à la remémoration inconscients des informations. L'apprentissage ou la récupération inconsciente d'informations par le biais de la mémoire procédurale , ou phénomène d'amorçage, en est un exemple . L'amorçage est le processus d' activation subliminale de réponses spécifiques issues de la mémoire et montre que toute la mémoire n'est pas activée consciemment, tandis que la mémoire procédurale est l'apprentissage lent et progressif de compétences qui se produit souvent sans attention consciente à l'apprentissage.
La mémoire n'est pas un système parfait et est affectée par de nombreux facteurs. Les mécanismes d'encodage, de stockage et de récupération de l'information peuvent tous être altérés. La douleur, par exemple, est un facteur physique qui altère la mémoire, comme l'ont montré des études sur des modèles animaux et des patients souffrant de douleurs chroniques. L'attention portée aux nouveaux stimuli peut réduire la quantité d'informations encodées pour le stockage. De plus, le processus de stockage peut être perturbé par des lésions physiques des zones cérébrales associées à la mémoire, telles que l'hippocampe. Enfin, la récupération d'informations en mémoire à long terme peut être affectée par la dégradation de cette mémoire. Le fonctionnement normal, la dégradation au fil du temps et les lésions cérébrales influencent tous la précision et la capacité de la mémoire.
Mémoire sensorielle
La mémoire sensorielle conserve les informations issues des sens moins d'une seconde après la perception d'un élément. La capacité à regarder un objet et à se souvenir de son apparence en une fraction de seconde d'observation, ou mémorisation, est un exemple de mémoire sensorielle. Elle échappe au contrôle cognitif et constitue une réponse automatique. Lors de présentations très courtes, les participants rapportent souvent avoir l'impression de « voir » plus d'éléments qu'ils ne peuvent en rapporter. Les premières expériences précises explorant cette forme de mémoire sensorielle ont été menées par George Sperling (1963) à l'aide du « paradigme du rapport partiel ». Les sujets étaient confrontés à une grille de 12 lettres, disposées en trois rangées de quatre. Après une brève présentation, un son aigu, médium ou grave leur était diffusé, leur indiquant la rangée à rapporter. Grâce à ces expériences de rapport partiel, Sperling a pu démontrer que la capacité de la mémoire sensorielle était d'environ 12 éléments, mais qu'elle diminuait très rapidement (en quelques centaines de millisecondes). Comme ce type de mémoire se dégrade très rapidement, les participants verraient l'affichage mais seraient incapables de restituer tous les éléments (12 dans la procédure de « rapport complet ») avant qu'ils ne disparaissent. Ce type de mémoire ne peut être prolongé par la répétition.
Il existe trois types de mémoire sensorielle. La mémoire iconique est une mémoire visuelle à dégradation rapide qui conserve brièvement une image perçue pendant une courte durée. La mémoire échoïque est une mémoire auditive à dégradation rapide qui conserve brièvement des sons perçus pendant une courte durée. La mémoire haptique est une mémoire sensorielle qui constitue une base de données pour les stimuli tactiles.
Indices de récupération
La manière dont nous encodons l'information a un impact considérable sur notre capacité à nous en souvenir ultérieurement. Les recherches sur les niveaux de traitement montrent que la mémoire est meilleure lorsque l'encodage privilégie les aspects « profonds » d'un stimulus (comme sa signification) plutôt que ses aspects « superficiels » (comme un lieu). Un encodage « profond » conduit à une représentation très distribuée, tandis qu'un encodage « superficiel » conduit à une représentation éparse. Se concentrer sur la signification permet de construire un souvenir plus riche et plus cohérent, tandis qu'un traitement superficiel laisse des traces plus faibles et moins durables. La mémoire fonctionne également mieux lorsque les indices utilisés pour se souvenir de l'information correspondent à ceux présents lors de l'apprentissage, ce qui est lié au principe de spécificité de l'encodage. De même, la dépendance au contexte montre que la mémorisation est facilitée lorsque l'environnement d'étude correspond à celui de l'évaluation. Par exemple, étudier dans un laboratoire peut être utile si l'évaluation se déroule dans le même environnement. En résumé, ces résultats montrent que la manière dont nous appréhendons l'information et le contexte dans lequel nous l'apprenons jouent un rôle majeur dans la mémoire.
mémoire à court terme
La mémoire à court terme, à ne pas confondre avec la mémoire de travail, permet de se souvenir d'informations pendant quelques secondes à une minute sans répétition. Sa capacité est cependant très limitée. En 1956, George A. Miller (1920-2012), alors qu'il travaillait aux Laboratoires Bell , a mené des expériences montrant que la mémoire à court terme pouvait contenir 7 ± 2 éléments (d'où le titre de son célèbre article : « Le nombre magique 7 ± 2 ») . Les études modernes estiment la capacité de la mémoire à court terme à un niveau inférieur, généralement de l'ordre de 4 à 5 éléments , ou proposent une limite plus flexible basée sur la quantité d'informations plutôt que sur le nombre d'éléments . La capacité de la mémoire peut être augmentée par un processus appelé segmentation en blocs (ou « chunking ») . Par exemple, pour se souvenir d'un numéro de téléphone à dix chiffres , on peut regrouper les chiffres en trois groupes : d'abord l'indicatif régional (comme le 123), puis un groupe de trois chiffres (456), et enfin un groupe de quatre chiffres (7890). Cette méthode de mémorisation est bien plus efficace que d'essayer de retenir une suite de dix chiffres ; en effet, elle permet de regrouper l'information en unités logiques. C'est pourquoi certains pays affichent les numéros de téléphone par groupes de deux à quatre chiffres.
On pense que la mémoire à court terme repose principalement sur un codage acoustique pour le stockage de l'information, et dans une moindre mesure sur un codage visuel. Conrad (1964) a constaté que les sujets testés avaient plus de difficultés à se souvenir de suites de lettres acoustiquement similaires, par exemple E, P, D. La confusion entre le rappel de lettres acoustiquement similaires et celui de lettres visuellement similaires suggère que les lettres étaient encodées acoustiquement. Cependant, l'étude de Conrad (1964) porte sur l'encodage de textes écrits. Ainsi, bien que la mémoire du langage écrit puisse reposer sur des composantes acoustiques, on ne peut généraliser ces résultats à toutes les formes de mémoire.
mémoire à long terme

La capacité et la durée de stockage de la mémoire sensorielle et de la mémoire à court terme sont généralement limitées. Autrement dit, l'information n'est pas conservée indéfiniment. En revanche, bien que la capacité totale de la mémoire à long terme reste à déterminer, elle peut stocker des quantités d'information bien plus importantes. De plus, elle peut conserver ces informations pendant une durée beaucoup plus longue, potentiellement toute une vie. Par exemple, face à un nombre aléatoire à sept chiffres, on peut ne s'en souvenir que quelques secondes avant de l'oublier, ce qui suggère qu'il a été stocké dans la mémoire à court terme. Par contre, on peut se souvenir des numéros de téléphone pendant de nombreuses années grâce à la répétition ; cette information est alors considérée comme stockée dans la mémoire à long terme.
Alors que la mémoire à court terme encode l'information de manière acoustique, la mémoire à long terme l'encode de manière sémantique : Baddeley (1966) a découvert qu'après 20 minutes, les sujets testés avaient le plus de difficultés à se souvenir à long terme d'une série de mots ayant des significations similaires (par exemple, grand, vaste, immense). Une autre composante de la mémoire à long terme est la mémoire épisodique, « qui tente de saisir des informations telles que le "quoi", le "quand" et le "où " » . Grâce à la mémoire épisodique, les individus sont capables de se souvenir d'événements précis tels que des fêtes d'anniversaire et des mariages.
La mémoire à court terme repose sur des schémas transitoires de communication neuronale, dépendant de régions du lobe frontal (notamment le cortex préfrontal dorsolatéral ) et du lobe pariétal . La mémoire à long terme, quant à elle, est maintenue par des modifications plus stables et permanentes des connexions neuronales, largement réparties dans le cerveau. L' hippocampe est essentiel (pour l'apprentissage de nouvelles informations) à la consolidation des informations de la mémoire à court terme vers la mémoire à long terme, bien qu'il ne semble pas stocker d'informations lui-même. On pensait que sans hippocampe, les nouveaux souvenirs ne pouvaient être stockés dans la mémoire à long terme et que la capacité d'attention serait très courte , comme l'ont initialement suggéré les observations du patient Henry Molaison après ce qui était considéré comme l'ablation complète de ses deux hippocampes. Un examen post-mortem plus récent de son cerveau montre que l'hippocampe était plus intact qu'on ne le pensait, remettant en question les théories tirées des données initiales. L'hippocampe peut être impliqué dans la modification des connexions neuronales pendant une période de trois mois ou plus après l'apprentissage initial.
Des recherches ont suggéré que le stockage de la mémoire à long terme chez l'homme pourrait être maintenu par la méthylation de l'ADN , et le gène « prion » .
Des recherches complémentaires ont exploré les bases moléculaires de la mémoire à long terme . Dès 2015, il était clair que la mémoire à long terme nécessite l'activation de la transcription génique et la synthèse de novo de protéines . La formation de la mémoire à long terme dépend à la fois de l'activation des gènes favorisant la mémoire et de l'inhibition des gènes suppresseurs de la mémoire, et la méthylation / déméthylation de l'ADN s'est révélée être un mécanisme majeur pour parvenir à cette double régulation
Chez les rats ayant acquis une nouvelle mémoire à long terme forte suite à un conditionnement de la peur contextuelle, l'expression d'environ 1 000 gènes est réduite tandis que celle d'environ 500 autres est augmentée dans l'hippocampe 24 heures après l'entraînement, ce qui correspond à une modification de l'expression de 9,17 % du génome hippocampique du rat. La réduction de l'expression génique est associée à la méthylation des gènes concernés.
Des recherches approfondies sur la mémoire à long terme ont permis d'élucider les mécanismes moléculaires d'établissement et de suppression des méthylations, comme décrit dans une revue de 2022 Ces mécanismes incluent, par exemple, les cassures double brin induites par TOP2B en réponse à un signal dans les gènes à expression précoce . De plus, les ARN messagers de nombreux gènes dont l'expression a été modulée par la méthylation sont transportés par les granules neuronaux ( RNP messagers ) jusqu'aux épines dendritiques . À ces niveaux, les ARN messagers peuvent être traduits en protéines qui contrôlent la signalisation au niveau des synapses neuronales .
Consolidation de la mémoire
La transition d'un souvenir de court terme à long terme est appelée consolidation mnésique . Les processus physiologiques impliqués restent mal connus. Deux hypothèses expliquent comment le cerveau accomplit cette tâche : la rétropropagation et la rétroaction positive du système endocrinien. La rétropropagation a été proposée comme mécanisme de consolidation mnésique et a notamment été utilisée par Geoffrey E. Hinton , prix Nobel de physique 2024, pour développer des logiciels d'intelligence artificielle. Elle suppose une rétroaction sur les neurones consolidant un souvenir donné, visant à effacer cette information lorsque le cerveau apprend qu'elle est trompeuse ou erronée. Cependant, aucune preuve empirique de son existence n'est disponible.
Au contraire, une rétroaction positive favorisant la consolidation d'une mémoire à court terme enregistrée dans les neurones et considérée comme utile par le système neuroendocrinien permettra à cette mémoire de se transformer en mémoire permanente. Ceci a été démontré expérimentalement chez les insectes été confirmée chez les oiseaux, les mammifères et d'autres créatures, y compris l'être humain
Les cellules gliales jouent également un rôle important dans la formation de la mémoire, bien que leur mode d'action reste à élucider.
D'autres mécanismes de consolidation de la mémoire ne peuvent être écartés.
Modèle multi-magasins

Le modèle multi-stockage (également connu sous le nom de modèle de mémoire d'Atkinson-Shiffrin ) a été décrit pour la première fois en 1968 par Atkinson et Shiffrin .
Le modèle multi-stockage a été critiqué pour son caractère simpliste. Par exemple, on pense que la mémoire à long terme est en réalité composée de plusieurs sous-composantes, comme la mémoire épisodique et la mémoire procédurale . Ce modèle postule également que la répétition est le seul mécanisme par lequel l'information atteint finalement le stockage à long terme, or des données montrent que nous sommes capables de nous souvenir de choses sans répétition.
Le modèle considère également l'ensemble des sources de mémoire comme une seule unité, alors que les recherches montrent des résultats différents. Par exemple, la mémoire à court terme peut être divisée en différentes unités, telles que les informations visuelles et auditives. Dans une étude de Zlonoga et Gerber (1986), le patient « KF » présentait certaines anomalies par rapport au modèle d'Atkinson-Shiffrin. Ce patient, atteint de lésions cérébrales , présentait des difficultés de mémoire à court terme. La reconnaissance des sons, tels que les chiffres, les lettres, les mots et les bruits facilement identifiables (comme les sonnettes et les miaulements de chat), était altérée. La mémoire visuelle à court terme, quant à elle, n'était pas affectée, ce qui suggère une dichotomie entre mémoire visuelle et auditive.
mémoire de travail

En 1974, Baddeley et Hitch ont proposé un « modèle de mémoire de travail » qui remplaçait le concept général de mémoire à court terme par le maintien actif de l’information dans un stockage à court terme. Dans ce modèle, la mémoire de travail se compose de trois systèmes de stockage de base : l’administrateur central, la boucle phonologique et le calepin visuo-spatial. En 2000, ce modèle a été enrichi par l’ajout du tampon épisodique multimodal ( modèle de mémoire de travail de Baddeley ).
L'organe exécutif central agit essentiellement comme un centre de stockage de l'attention. Il canalise les informations vers les trois processus qui le composent : la boucle phonologique, le calepin visuo-spatial et la mémoire épisodique.
La boucle phonologique stocke l'information auditive en répétant silencieusement des sons ou des mots de manière continue : c'est le processus articulatoire (par exemple, la répétition d'un numéro de téléphone). Une courte liste d'informations est plus facile à mémoriser. La boucle phonologique est parfois perturbée. Des paroles parasites ou des bruits de fond peuvent l'entraver. La suppression articulatoire peut également perturber l'encodage, et des mots à la sonorité proche peuvent être intervertis ou mal mémorisés en raison de l'effet de similarité phonologique. La boucle phonologique a également une capacité de stockage limitée, ce qui explique qu'il soit plus facile de se souvenir de nombreux mots courts que de nombreux mots longs, conformément à l'effet de longueur des mots.
Le calepin visuo-spatial stocke les informations visuelles et spatiales. Il est sollicité lors de la réalisation de tâches spatiales (comme l'estimation des distances) ou visuelles (comme le comptage des fenêtres d'une maison ou la visualisation d'images). Les personnes atteintes d' aphantasie ne peuvent pas utiliser le calepin visuo-spatial.
La mémoire tampon épisodique sert à relier les informations provenant de différents domaines afin de former des unités intégrées d'informations visuelles, spatiales et verbales, ordonnées chronologiquement (par exemple, le souvenir d'une histoire ou d'une scène de film). On suppose également que la mémoire tampon épisodique est liée à la mémoire à long terme et au sens sémantique.
Le modèle de la mémoire de travail explique de nombreuses observations pratiques, notamment pourquoi il est plus facile d'effectuer deux tâches différentes, l'une verbale et l'autre visuelle, que deux tâches similaires, et l'effet de longueur des mots mentionné précédemment. La mémoire de travail est également à la base de ce qui nous permet d'accomplir les activités quotidiennes impliquant la pensée. C'est la partie de la mémoire où nous menons les processus de pensée et les utilisons pour apprendre et raisonner sur divers sujets.
Types
Les chercheurs font la distinction entre la mémoire de reconnaissance et la mémoire de rappel . Les tâches de reconnaissance consistent à demander aux individus d'indiquer s'ils ont déjà rencontré un stimulus (comme une image ou un mot). Les tâches de rappel consistent à demander aux participants de récupérer des informations apprises antérieurement. Par exemple, on peut leur demander de reproduire une série d'actions qu'ils ont déjà vues ou de réciter une liste de mots qu'ils ont déjà entendus.
Par type d'information
La mémoire topographique implique la capacité de s'orienter dans l'espace, de reconnaître et de suivre un itinéraire, ou de reconnaître des lieux familiers. Se perdre en voyageant seul est un exemple de défaillance de la mémoire topographique.
Les souvenirs flash sont des souvenirs épisodiques clairs d'événements uniques et très émotionnels. Les personnes qui se souviennent de ce qu'elles faisaient lorsqu'elles ont appris la nouvelle de l' assassinat du président Kennedy , des attentats du 11 septembre ou du siège de Sydney sont des exemples de souvenirs flash.
à long terme
Anderson (1976) divise la mémoire à long terme en mémoire déclarative (explicite) et en mémoire procédurale (implicite) .
Déclaratif
La mémoire déclarative requiert un rappel conscient , c'est-à-dire qu'un processus conscient doit faire remonter l'information. On l'appelle parfois mémoire explicite , puisqu'elle consiste en des informations stockées et récupérées explicitement. La mémoire déclarative se subdivise en mémoire sémantique , qui concerne les principes et les faits indépendamment du contexte, et en mémoire épisodique , qui concerne les informations spécifiques à un contexte particulier, comme un lieu et un moment précis. La mémoire sémantique permet d'encoder des connaissances abstraites sur le monde, comme « Paris est la capitale de la France ». La mémoire épisodique, quant à elle, est utilisée pour des souvenirs plus personnels, tels que les sensations, les émotions et les associations personnelles liées à un lieu ou un moment précis. Les souvenirs épisodiques reflètent souvent les « premières fois » de la vie, comme un premier baiser, un premier jour d'école ou une première victoire en championnat. Ce sont des événements marquants de la vie dont on se souvient clairement.
Des recherches suggèrent que la mémoire déclarative repose sur plusieurs fonctions du lobe temporal médian, notamment l'hippocampe. La mémoire autobiographique – le souvenir d'événements particuliers de sa propre vie – est généralement considérée comme équivalente à la mémoire épisodique, ou comme un sous-ensemble de celle-ci. La mémoire visuelle fait partie de la mémoire qui préserve certaines caractéristiques de nos sens liées à l'expérience visuelle. On est capable de mémoriser des informations qui ressemblent à des objets, des lieux, des animaux ou des personnes, sous forme d' images mentales . La mémoire visuelle peut induire un effet d'amorçage , et l'on suppose qu'un système de représentation perceptive sous-tend ce phénomène.
De procédure
À l'inverse, la mémoire procédurale (ou mémoire implicite ) ne repose pas sur le rappel conscient d'informations, mais sur un apprentissage implicite . On peut la résumer comme la capacité à se souvenir comment faire quelque chose. La mémoire procédurale est principalement utilisée dans l'apprentissage des habiletés motrices et peut être considérée comme un sous-ensemble de la mémoire implicite. Elle se manifeste lorsqu'une personne réussit mieux une tâche donnée grâce à la simple répétition : aucun nouveau souvenir explicite n'a été formé, mais on accède inconsciemment à des aspects de ces expériences antérieures. La mémoire procédurale impliquée dans l'apprentissage moteur dépend du cervelet et des ganglions de la base .
Une caractéristique de la mémoire procédurale est que les éléments mémorisés sont automatiquement traduits en actions, et sont donc parfois difficiles à décrire. La capacité à faire du vélo ou à lacer ses chaussures en est un exemple.
Par direction temporelle
Une autre distinction majeure entre les différentes fonctions de la mémoire réside dans la nature du contenu à se souvenir : passé ( mémoire rétrospective ) ou futur ( mémoire prospective ). John Meacham a introduit cette distinction lors d'une communication présentée au congrès annuel de l'American Psychological Association en 1975 , puis reprise par Ulric Neisser dans son ouvrage collectif de 1982, *Memory Observed: Remembering in Natural Contexts . Ainsi, la mémoire rétrospective englobe la mémoire sémantique, épisodique et autobiographique. La mémoire prospective, quant à elle, concerne les intentions futures, ou le fait de se souvenir de se souvenir (Winograd, 1988). Elle se subdivise en deux catégories : la mémoire prospective événementielle et la mémoire prospective temporelle. Les souvenirs prospectifs temporels sont déclenchés par un repère temporel, comme aller chez le médecin (action) à 16 h (repère). Les souvenirs prospectifs événementiels sont des intentions déclenchées par des repères, comme se souvenir de poster une lettre (action) après avoir vu une boîte aux lettres (repère). Les indices n'ont pas besoin d'être liés à l'action (comme dans l'exemple de la boîte aux lettres/lettre), et les listes, les notes autocollantes, les mouchoirs noués ou la ficelle autour du doigt sont autant d'exemples d'indices que les gens utilisent comme stratégies pour améliorer leur mémoire prospective.
Techniques d'étude
Pour évaluer les nourrissons
Les nourrissons ne possèdent pas les capacités linguistiques nécessaires pour exprimer leurs souvenirs ; par conséquent, les récits verbaux ne peuvent servir à évaluer la mémoire des très jeunes enfants. Au fil des années, les chercheurs ont toutefois adapté et développé plusieurs outils d'évaluation de la mémoire de reconnaissance et de la mémoire de rappel chez les nourrissons. Les techniques d'habituation et de conditionnement opérant ont été utilisées pour évaluer la mémoire de reconnaissance, tandis que les techniques d'imitation différée et provoquée ont permis d'évaluer la mémoire de rappel.
Les techniques utilisées pour évaluer la mémoire de reconnaissance des nourrissons comprennent les suivantes :
- Procédure de comparaison visuelle par paires (basée sur l'habituation) : on présente d'abord aux nourrissons des paires de stimuli visuels, par exemple deux photos en noir et blanc de visages humains, pendant une durée déterminée ; puis, après s'être familiarisés avec les deux photos, on leur présente la photo « familière » et une nouvelle photo. Le temps passé à regarder chaque photo est enregistré. Un temps passé à regarder plus longtemps la nouvelle photo indique qu'ils se souviennent de la photo « familière ». Des études utilisant cette procédure ont montré que les nourrissons de 5 à 6 mois peuvent retenir des informations pendant quatorze jours.
- Technique de conditionnement opérant : les nourrissons sont placés dans un berceau et un ruban relié à un mobile suspendu au-dessus d’eux est attaché à l’un de leurs pieds. Les nourrissons constatent que lorsqu’ils donnent des coups de pied, le mobile se met en mouvement ; la fréquence de ces mouvements augmente considérablement en quelques minutes. Des études utilisant cette technique ont révélé que la mémoire des nourrissons s’améliore sensiblement au cours des 18 premiers mois. Alors que les bébés de 2 à 3 mois peuvent conserver une réponse opérante (comme l’activation du mobile par un coup de pied) pendant une semaine, ceux de 6 mois peuvent la conserver pendant deux semaines et ceux de 18 mois peuvent conserver une réponse opérante similaire jusqu’à 13 semaines.
Les techniques utilisées pour évaluer la mémoire de rappel des nourrissons comprennent les suivantes :
- Technique d’imitation différée : un expérimentateur montre à des nourrissons une séquence d’actions unique (par exemple, utiliser un bâtonnet pour appuyer sur un bouton d’une boîte) puis, après un délai, leur demande d’imiter ces actions. Des études utilisant l’imitation différée ont montré que la mémorisation de cette séquence d’actions par des enfants de 14 mois peut durer jusqu’à quatre mois.
- La technique d'imitation provoquée est très similaire à la technique d'imitation différée ; la différence réside dans le fait que les nourrissons sont autorisés à imiter les actions avant le délai. Des études utilisant la technique d'imitation provoquée ont montré que des enfants de 20 mois peuvent se rappeler les séquences d'actions douze mois plus tard.
Pour évaluer les enfants et les personnes âgées
Les chercheurs utilisent diverses tâches pour évaluer la mémoire des enfants plus âgés et des adultes. En voici quelques exemples :
- L’apprentissage par association de paires consiste à apprendre à associer un mot spécifique à un autre. Par exemple, lorsqu’on lui donne le mot « sûr », il doit apprendre à dire un autre mot spécifique, comme « vert ». Il s’agit d’un apprentissage par stimulus et réponse.
- Rappel libre – lors de cette tâche, le sujet est invité à étudier une liste de mots, puis à se souvenir ou à écrire le plus grand nombre de mots possible, comme pour les questions à réponse libre. Les premiers éléments présentés sont affectés par l'interférence rétroactive (IR), ce qui signifie que plus la liste est longue, plus l'interférence est importante et moins les mots ont de chances d'être rappelés. En revanche, les éléments présentés en dernier subissent peu d'IR, mais beaucoup d'interférence proactive (IP), ce qui signifie que plus le délai de rappel est long, plus les mots risquent d'être oubliés.
- Rappel indicé : on donne à la personne des indices significatifs pour l’aider à récupérer des informations préalablement encodées dans sa mémoire ; il peut s’agir généralement d’un mot en rapport avec l’information à rappeler. Cette méthode est similaire aux exercices à trous utilisés en classe.
- Reconnaissance – On demande aux sujets de se souvenir d’une liste de mots ou d’images, puis d’identifier les mots ou images présentés précédemment parmi une liste d’alternatives qui ne figuraient pas dans la liste originale. Ceci est similaire aux évaluations à choix multiples.
- Paradigme de détection : on présente aux participants un certain nombre d’objets et d’échantillons de couleurs pendant une période donnée. On teste ensuite leur capacité visuelle à mémoriser le plus d’éléments possible en leur demandant d’observer des images et d’indiquer si celles-ci sont similaires à l’échantillon ou si une différence est présente.
- La méthode des économies compare la vitesse d'apprentissage initiale à la vitesse de réapprentissage. Le temps économisé mesure la mémoire.
- Tâches de mémoire implicite – l'information est puisée en mémoire sans prise de conscience.
Échecs

- La fugacité des souvenirs – ou altération de la mémoire – se manifeste lors de la phase de stockage, entre l'encodage et la récupération de l'information. Ce phénomène peut concerner la mémoire sensorielle, la mémoire à court terme et la mémoire à long terme. De manière générale, l'information est rapidement oubliée durant les premiers jours ou les premières années, puis sa perte diminue progressivement au fil du temps.
- Distraction – Troubles de la mémoire dus à un manque d' attention . L'attention joue un rôle essentiel dans la mémorisation à long terme ; sans une attention suffisante, l'information risque de ne pas être stockée et, par conséquent, impossible à récupérer ultérieurement.
Physiologie
Les régions cérébrales impliquées dans la neuroanatomie de la mémoire, telles que l' hippocampe , l' amygdale , le striatum ou les corps mamillaires , seraient impliquées dans des types spécifiques de mémoire. Par exemple, l'hippocampe serait impliqué dans l'apprentissage spatial et l'apprentissage déclaratif , tandis que l'amygdale serait impliquée dans la mémoire émotionnelle .
Les lésions de certaines zones chez les patients et les modèles animaux, et les déficits de mémoire qui en résultent, constituent une source d'information primordiale. Cependant, plutôt que d'incriminer une zone spécifique, il se pourrait que des lésions de zones adjacentes, ou d'une voie traversant cette zone, soient en réalité responsables du déficit observé. De plus, il est insuffisant de décrire la mémoire, et son corollaire, l'apprentissage , comme dépendant uniquement de régions cérébrales spécifiques. L'apprentissage et la mémoire sont généralement attribués à des modifications des synapses neuronales , que l'on pense être médiées par la potentialisation à long terme (PLT) et la dépression à long terme (DLT) .
En général, plus un événement ou une expérience est chargé(e) émotionnellement, mieux on s'en souvient ; ce phénomène est connu sous le nom d' effet d'amélioration de la mémoire . Cependant, les patients présentant des lésions de l'amygdale ne manifestent pas cet effet.
Hebb distinguait la mémoire à court terme de la mémoire à long terme. Il postulait que tout souvenir maintenu en mémoire à court terme pendant une période suffisamment longue serait consolidé en mémoire à long terme. Des recherches ultérieures ont démontré que cette hypothèse était fausse. Il a été montré que des injections directes de cortisol ou d'adrénaline favorisent la mémorisation des expériences récentes. Ceci est également vrai pour la stimulation de l'amygdale. Cela prouve que l'excitation améliore la mémoire en stimulant la production d'hormones qui agissent sur l'amygdale. Un stress excessif ou prolongé (avec un taux de cortisol prolongé) peut nuire à la mémorisation. Les patients présentant des lésions de l'amygdale ne sont pas plus susceptibles de se souvenir de mots chargés émotionnellement que de mots non chargés émotionnellement. L'hippocampe est important pour la mémoire explicite. Il est également important pour la consolidation de la mémoire. L'hippocampe reçoit des informations de différentes parties du cortex et envoie ses efférences à différentes parties du cerveau. Ces informations proviennent des aires sensorielles secondaires et tertiaires qui les ont déjà largement traitées. Des lésions de l'hippocampe peuvent également entraîner des pertes de mémoire et des problèmes de mémorisation. Cette perte de mémoire comprend l'amnésie rétrograde , qui est la perte de mémoire des événements survenus peu de temps avant la lésion cérébrale.
Neurosciences cognitives
Les neuroscientifiques cognitifs définissent la mémoire comme la rétention, la réactivation et la reconstruction d'une représentation interne indépendante de l'expérience. Le terme de représentation interne implique que cette définition de la mémoire comporte deux composantes : l'expression de la mémoire au niveau comportemental ou conscient, et les modifications neuronales physiques sous-jacentes (Dudai, 2007). Cette dernière composante est également appelée engramme ou trace mnésique (Semon, 1904). Certains neuroscientifiques et psychologues assimilent à tort les concepts d'engramme et de mémoire, considérant de manière générale comme mémoire l'ensemble des séquelles persistantes des expériences ; d'autres s'opposent à cette idée, arguant que la mémoire n'existe que lorsqu'elle se manifeste dans le comportement ou la pensée (Moscovitch, 2007).
Une question cruciale en neurosciences cognitives est celle du codage et de la représentation de l'information et des expériences mentales dans le cerveau. Les études sur la plasticité neuronale ont permis d'acquérir de nombreuses connaissances sur les codes neuronaux, mais la plupart de ces recherches se sont concentrées sur l'apprentissage simple au sein de circuits neuronaux simples. Les modifications neuronales impliquées dans des formes de mémoire plus complexes, notamment la mémoire déclarative qui requiert le stockage de faits et d'événements, restent beaucoup moins claires (Byrne, 2007). Les zones de convergence-divergence pourraient constituer les réseaux neuronaux où les souvenirs sont stockés et récupérés. Étant donné la diversité des types de mémoire, qui dépendent des types de connaissances représentées, des mécanismes sous-jacents, des processus et des modes d'acquisition, il est probable que différentes aires cérébrales prennent en charge différents systèmes de mémoire et qu'elles soient interdépendantes au sein des réseaux neuronaux : « les composantes de la représentation mnésique sont largement distribuées dans différentes parties du cerveau, via de multiples circuits néocorticaux »
- L’ encodage de la mémoire de travail implique l’émission de potentiels d’action par des neurones individuels, induite par un stimulus sensoriel et qui persiste même après la disparition de ce stimulus (Jensen et Lisman, 2005 ; Fransen et al., 2002). L’encodage de la mémoire épisodique implique des modifications persistantes des structures moléculaires qui altèrent la transmission synaptique entre les neurones. Parmi ces modifications structurales, on peut citer la potentialisation à long terme (PLT) ou la plasticité synaptique dépendante du temps d’émission (PST). L’activité neuronale persistante en mémoire de travail peut amplifier les modifications synaptiques et cellulaires lors de l’encodage de la mémoire épisodique (Jensen et Lisman, 2005).
- Mémoire de travail. Des études d'imagerie fonctionnelle récentes ont détecté des signaux de mémoire de travail à la fois dans le lobe temporal médian (LTM), une région cérébrale fortement associée à la mémoire à long terme , et dans le cortex préfrontal (Ranganath et al., 2005), suggérant une forte corrélation entre la mémoire de travail et la mémoire à long terme. Cependant, l'abondance significative de signaux de mémoire de travail observés dans le lobe préfrontal suggère que cette région joue un rôle plus important dans la mémoire de travail que le LTM (Suzuki, 2007).
- Consolidation et reconsolidation . La mémoire à court terme (MCT) est temporaire et sujette à des perturbations, tandis que la mémoire à long terme (MLT), une fois consolidée, est persistante et stable. La consolidation de la MCT en MLT au niveau moléculaire impliquerait deux processus : la consolidation synaptique et la consolidation systémique. La première fait intervenir la synthèse protéique dans le lobe temporal médian (LTM), tandis que le second transforme la mémoire dépendante du LTM en une mémoire indépendante du LTM sur une période allant de plusieurs mois à plusieurs années (Ledoux, 2007). Ces dernières années, ce dogme traditionnel de la consolidation a été réévalué à la lumière des études sur la reconsolidation. Ces études ont montré que la prévention après la récupération influence la récupération ultérieure du souvenir (Sara, 2000). De nouvelles études ont démontré qu’un traitement post-récupération par des inhibiteurs de la synthèse protéique et de nombreux autres composés peut induire un état amnésique (Nadel et al., 2000b ; Alberini, 2005 ; Dudai, 2006). Ces résultats sur la reconsolidation concordent avec les données comportementales montrant que les souvenirs récupérés ne sont pas une copie conforme des expériences initiales, et que les souvenirs sont mis à jour lors de leur récupération.
Génétique
L'étude de la génétique de la mémoire humaine n'en est qu'à ses débuts, bien que de nombreux gènes aient été étudiés pour leur association avec la mémoire chez l'homme et l'animal. Un succès initial notable a été l'association du gène APOE avec les troubles de la mémoire dans la maladie d'Alzheimer . La recherche de gènes associés à la variabilité normale de la mémoire se poursuit. L'un des premiers candidats pour expliquer cette variabilité est la protéine KIBRA [ qui semble être associée à la vitesse à laquelle les informations sont oubliées au fil du temps. Certaines données suggèrent que les souvenirs sont stockés dans le noyau des neurones
Bases génétiques
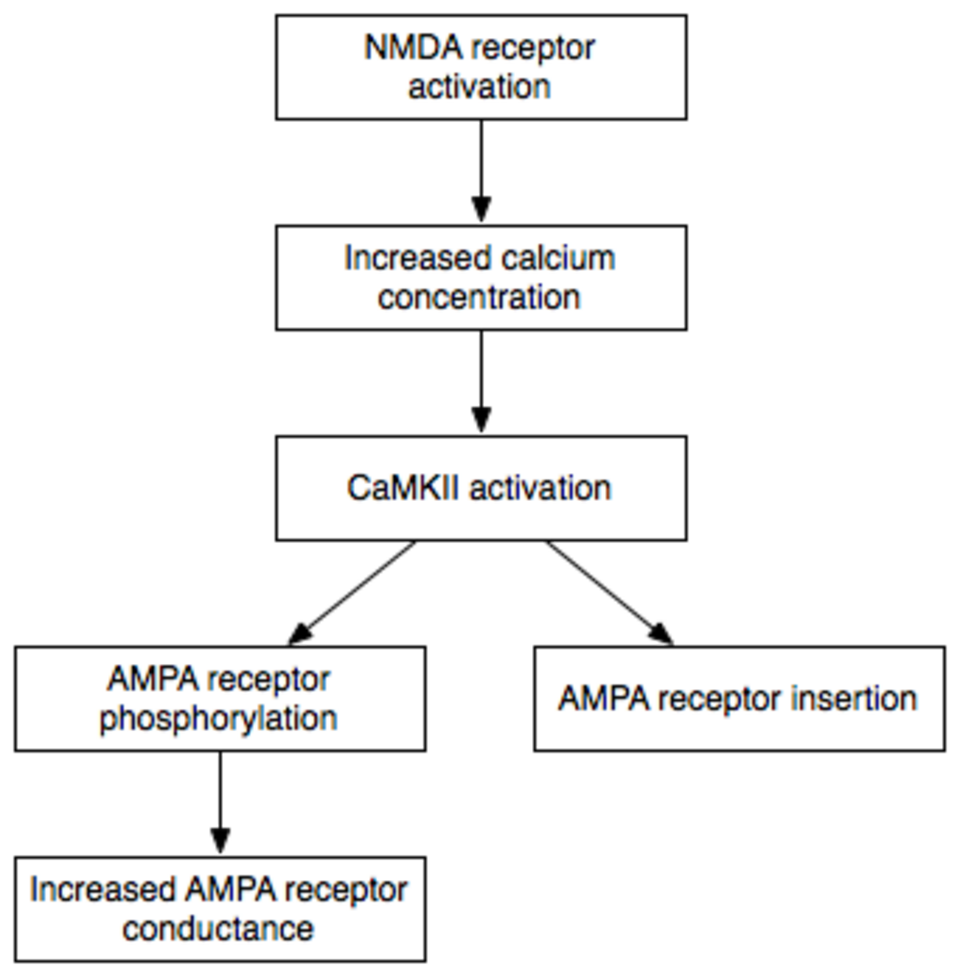
Plusieurs gènes , protéines et enzymes ont fait l'objet de recherches approfondies concernant leur implication dans la mémoire. La mémoire à long terme, contrairement à la mémoire à court terme, dépend de la synthèse de nouvelles protéines . Ce processus se déroule au sein de la cellule et concerne les neurotransmetteurs, les récepteurs et les nouvelles voies synaptiques qui renforcent la communication entre les neurones. La production de nouvelles protéines dédiées au renforcement synaptique est déclenchée par la libération de certaines substances de signalisation (comme le calcium dans les neurones de l'hippocampe). Dans le cas des cellules hippocampiques, cette libération dépend de l'expulsion du magnésium (une molécule de liaison) qui se produit après une signalisation synaptique importante et répétée. L'expulsion temporaire du magnésium libère les récepteurs NMDA , ce qui entraîne la libération de calcium dans la cellule. Ce signal induit la transcription de gènes et la synthèse de protéines de renforcement . Pour plus d'informations, voir la potentialisation à long terme (PLT).
L'une des protéines nouvellement synthétisées lors de la potentialisation à long terme (LTP) est également essentielle au maintien de la mémoire à long terme. Il s'agit d'une forme active autonome de l'enzyme protéine kinase C (PKC), appelée PKMζ . La PKMζ maintient le renforcement synaptique dépendant de l'activité neuronale et son inhibition efface les souvenirs à long terme établis, sans affecter la mémoire à court terme. De plus, une fois l'inhibiteur éliminé, la capacité d'encoder et de stocker de nouveaux souvenirs à long terme est restaurée. Le BDNF est également important pour la persistance des souvenirs à long terme.
La stabilisation à long terme des modifications synaptiques est également déterminée par une augmentation parallèle des structures pré- et postsynaptiques, telles que les boutons axonaux , les épines dendritiques et la densité postsynaptique . Au niveau moléculaire, une augmentation des protéines d'échafaudage postsynaptiques PSD-95 et HOMER1c est corrélée à la stabilisation de l'élargissement synaptique . La protéine de liaison à l'élément de réponse à l'AMPc ( CREB ) est un facteur de transcription qui jouerait un rôle important dans la consolidation des souvenirs à court terme en souvenirs à long terme et dont l'expression serait diminuée dans la maladie d'Alzheimer
Méthylation et déméthylation de l'ADN

Les rats exposés à un apprentissage intense peuvent en conserver un souvenir permanent, même après une seule séance. Ce souvenir à long terme semble initialement stocké dans l' hippocampe , mais ce stockage est transitoire. Une grande partie de ce stockage semble se dérouler dans le cortex cingulaire antérieur . Lors d'une telle exposition expérimentale, plus de 5 000 régions d'ADN différemment méthylées sont apparues dans le génome neuronal de l'hippocampe des rats, une heure et 24 heures après l'entraînement [94]. profil de méthylation ont affecté de nombreux gènes sous -exprimés , souvent en raison de la formation de nouveaux sites de 5-méthylcytosine dans des régions riches en CpG du génome. Par ailleurs, de nombreux autres gènes ont été surexprimés, probablement souvent par hypométhylation. L'hypométhylation résulte fréquemment de la suppression de groupements méthyle sur des 5-méthylcytosines préexistantes dans l'ADN. La déméthylation est réalisée par plusieurs protéines agissant de concert, notamment les enzymes TET ainsi que les enzymes de la voie de réparation par excision de base de l'ADN (voir Épigénétique et apprentissage ). Le profil d'expression des gènes induits et réprimés dans les neurones cérébraux à la suite d'un apprentissage intense constitue probablement la base moléculaire de la mémorisation à long terme de cet événement.
Épigénétique
Les études sur les bases moléculaires de la formation de la mémoire indiquent que les mécanismes épigénétiques agissant dans les neurones cérébraux jouent un rôle central dans la détermination de cette capacité. Parmi les principaux mécanismes épigénétiques impliqués dans la mémoire figurent la méthylation et la déméthylation de l'ADN neuronal, ainsi que les modifications des protéines histones , notamment les méthylations , les acétylations et les désacétylations .
La stimulation de l'activité cérébrale lors de la formation de la mémoire s'accompagne souvent de lésions de l'ADN neuronal , suivies d'une réparation associée à des modifications épigénétiques persistantes. En particulier, les processus de réparation de l'ADN par jonction d'extrémités non homologues et par excision de base sont impliqués dans la formation de la mémoire.
L'ADN topoisomérase 2-bêta dans l'apprentissage et la mémoire
Lors d'un nouvel apprentissage, un ensemble de gènes s'exprime rapidement dans le cerveau. Cette expression génique induite est considérée comme essentielle au traitement de l'information apprise. Ces gènes sont appelés gènes à expression précoce (IEG). L'activité de la topoisomérase 2-bêta (TOP2B) est essentielle à l'expression des IEG lors d'un type d'apprentissage chez la souris appelé mémoire associative de la peur . Ce type d'apprentissage semble déclencher rapidement l'activité de TOP2B, induisant des cassures double brin dans l' ADN promoteur des gènes IEG impliqués dans la neuroplasticité . La réparation de ces cassures induites est associée à la déméthylation de l'ADN des promoteurs des gènes IEG, permettant ainsi l'expression immédiate de ces gènes
Les cassures double brin induites lors d'un apprentissage ne sont pas immédiatement réparées. Environ 600 séquences régulatrices dans les promoteurs et environ 800 séquences régulatrices dans les activateurs semblent dépendre des cassures double brin initiées par la topoisomérase 2-bêta (TOP2B) pour leur activation. L'induction de certaines cassures double brin est spécifique à leur signal inducteur. Lors de l'activation de neurones in vitro , seules 22 cassures double brin induites par TOP2B sont observées dans leur génome.
Ces cassures double brin induites par TOP2B s'accompagnent de l'intervention d'au moins quatre enzymes de la voie de réparation de l'ADN par jonction d'extrémités non homologues (NHEJ) (DNA-PKcs, KU70, KU80 et ADN ligase IV) (voir figure). Ces enzymes réparent les cassures double brin en 15 minutes à deux heures environ. Les cassures double brin dans le promoteur sont donc associées à TOP2B et à au moins ces quatre enzymes de réparation. Ces protéines sont présentes simultanément sur un seul nucléosome promoteur (environ 147 nucléotides de la séquence d'ADN s'enroulent autour d'un nucléosome) situé près du site d'initiation de la transcription de leur gène cible.

La cassure double brin induite par TOP2B libère apparemment la partie du promoteur située au niveau du site d'initiation de la transcription lié à l'ARN polymérase , lui permettant ainsi de se déplacer physiquement vers son activateur associé (voir la séquence régulatrice ). Ceci permet à l'activateur, avec ses facteurs de transcription et protéines médiatrices liés , d'interagir directement avec l'ARN polymérase bloquée au niveau du site d'initiation de la transcription afin de déclencher la transcription .
Le conditionnement de la peur contextuelle chez la souris induit une mémorisation à long terme et une peur du lieu où l'événement s'est produit. Ce conditionnement provoque des centaines de cassures double brin (CDB) dans les neurones du cortex préfrontal médian (mPFC) et de l'hippocampe (voir Figure : Régions cérébrales impliquées dans la formation de la mémoire). Ces CDB activent principalement des gènes impliqués dans les processus synaptiques, essentiels à l'apprentissage et à la mémoire.
Dans la petite enfance
Jusqu'au milieu des années 1980, on supposait que les nourrissons étaient incapables d'encoder, de retenir et de restituer des informations. De plus en plus d'études indiquent que des nourrissons dès l'âge de 6 mois peuvent se souvenir d'informations après un délai de 24 heures. Par ailleurs, les recherches ont révélé qu'avec l'âge, les nourrissons peuvent stocker des informations pendant des périodes plus longues : les enfants de 6 mois peuvent se souvenir d'informations après 24 heures, ceux de 9 mois après cinq semaines et ceux de 20 mois après douze mois. En outre, des études ont montré qu'avec l'âge, les nourrissons stockent les informations plus rapidement. Alors que les enfants de 14 mois peuvent se souvenir d'une séquence de trois étapes après une seule exposition, les enfants de 6 mois ont besoin d'environ six expositions pour pouvoir la mémoriser.
Bien que les bébés de six mois puissent se souvenir d'informations à court terme, ils ont des difficultés à se rappeler l'ordre temporel de ces informations. Ce n'est qu'à neuf mois que les nourrissons sont capables de se rappeler les actions d'une séquence en deux étapes dans le bon ordre chronologique, c'est-à-dire l'étape 1 puis l'étape 2. Autrement dit, lorsqu'on leur demande d'imiter une séquence d'actions en deux étapes (comme placer une petite voiture dans sa base et appuyer sur le piston pour la faire rouler jusqu'à l'autre extrémité), les bébés de neuf mois ont tendance à imiter les actions de la séquence dans le bon ordre (étape 1 puis étape 2). Les nourrissons plus jeunes (6 mois) ne peuvent se rappeler qu'une seule étape d'une séquence en deux étapes. Les chercheurs ont suggéré que ces différences liées à l'âge sont probablement dues au fait que le gyrus denté de l'hippocampe et les composantes frontales du réseau neuronal ne sont pas complètement développés à l'âge de six mois.
En réalité, le terme « amnésie infantile » désigne le phénomène d’oubli accéléré durant la petite enfance. Il est important de noter que l’amnésie infantile n’est pas propre à l’espèce humaine, et les recherches précliniques (utilisant des modèles rongeurs) permettent de mieux comprendre la neurobiologie précise de ce phénomène. Une analyse de la littérature réalisée par le neuroscientifique comportementaliste Jee Hyun Kim suggère que l’oubli accéléré en début de vie est dû, au moins en partie, à la croissance rapide du cerveau durant cette période.
Vieillissement
L'une des principales préoccupations des personnes âgées est la perte de mémoire, d'autant plus qu'il s'agit d'un symptôme caractéristique de la maladie d'Alzheimer . Cependant, la perte de mémoire liée au vieillissement normal diffère qualitativement de celle associée à un diagnostic de maladie d'Alzheimer (Budson & Price, 2005). Des recherches ont révélé que les performances des individus aux tâches de mémoire sollicitant les régions frontales diminuent avec l'âge. Les personnes âgées ont tendance à présenter des déficits aux tâches impliquant la connaissance de l'ordre chronologique de l'apprentissage d'une information , aux tâches de mémoire source qui exigent de se souvenir des circonstances ou du contexte précis de cet apprentissage , et aux tâches de mémoire prospective qui consistent à se souvenir d'une action à effectuer ultérieurement. L'utilisation d'un agenda, par exemple, peut aider les personnes âgées à gérer leurs difficultés de mémoire prospective.
Les profils de transcription génique ont été déterminés dans le cortex frontal humain d'individus âgés de 26 à 106 ans. De nombreux gènes ont présenté une expression réduite après 40 ans, et particulièrement après 70 ans Parmi ceux présentant la réduction d'expression la plus significative avec l'âge figuraient des gènes jouant un rôle central dans la mémoire et l'apprentissage . On a également observé une augmentation marquée des dommages à l'ADN , probablement des dommages oxydatifs , dans les promoteurs de ces gènes à expression réduite. Il a été suggéré que les dommages à l'ADN pourraient réduire l'expression de gènes particulièrement vulnérables impliqués dans la mémoire et l'apprentissage
Troubles
Une grande partie des connaissances actuelles sur la mémoire provient de l'étude des troubles de la mémoire , en particulier la perte de mémoire, appelée amnésie . L'amnésie peut résulter de lésions importantes : (a) des régions du lobe temporal médian, telles que l'hippocampe, le gyrus denté, le subiculum, l'amygdale, les cortex parahippocampique, entorhinal et périrhinal ou (b) de la région diencéphalique médiane, plus précisément du noyau dorsomédian du thalamus et des corps mamillaires de l'hypothalamus . Il existe de nombreux types d'amnésie, et l'étude de leurs différentes formes a permis d'observer des anomalies apparentes dans les sous-systèmes cérébraux de la mémoire, et ainsi de formuler des hypothèses sur leur fonction dans un cerveau sain. D'autres troubles neurologiques, comme la maladie d'Alzheimer et la maladie de Parkinson peuvent également affecter la mémoire et la cognition. L’hyperthymésie , ou syndrome hyperthymésique, est un trouble qui affecte la mémoire autobiographique d’un individu, ce qui signifie essentiellement qu’il ne peut oublier de petits détails qui, autrement, ne seraient pas stockés. Le syndrome de Korsakoff , également connu sous le nom de psychose de Korsakoff ou syndrome amnésique-confabulatoire, est une maladie cérébrale organique qui affecte la mémoire par une perte ou une réduction généralisée des neurones dans le cortex préfrontal.
Bien qu'il ne s'agisse pas d'un trouble, le phénomène du « mot sur le bout de la langue » est une difficulté passagère et fréquente à retrouver ses mots en mémoire. Cependant, les personnes atteintes d'aphasie anomique (également appelée aphasie nominale ou anomie) présentent ce phénomène de manière permanente en raison de lésions des lobes frontaux et pariétaux du cerveau .
Des troubles de la mémoire peuvent également survenir après une infection virale. De nombreux patients convalescents de la COVID-19 présentent des pertes de mémoire . D'autres virus peuvent également provoquer des troubles de la mémoire, notamment le SARS-CoV-1 , le MERS-CoV , le virus Ebola et même le virus de la grippe .
Facteurs d'influence
Ingérence
L'interférence peut entraver la mémorisation et la restitution des connaissances. On distingue l'interférence rétroactive , lorsque l'apprentissage de nouvelles informations rend plus difficile le rappel d'informations antérieures et l'interférence proactive , où les apprentissages antérieurs perturbent la restitution de nouvelles informations. Bien que l'interférence puisse entraîner l'oubli, il est important de garder à l'esprit que, dans certaines situations, les connaissances antérieures peuvent faciliter l'apprentissage de nouvelles informations. Par exemple, la connaissance du latin peut aider à apprendre une langue apparentée comme le français ; ce phénomène est connu sous le nom de transfert positif
Stresser
Le stress a un impact significatif sur la formation de la mémoire et l'apprentissage. En réponse à des situations stressantes, le cerveau libère des hormones et des neurotransmetteurs (par exemple, des glucocorticoïdes et des catécholamines) qui affectent les processus d'encodage de la mémoire dans l'hippocampe. Des études comportementales sur des animaux montrent que le stress chronique induit la production d'hormones surrénaliennes qui influencent la structure de l'hippocampe chez le rat . Une étude expérimentale menée par les psychologues cognitifs allemands L. Schwabe et O. Wolf démontre que l'apprentissage sous stress diminue également la capacité de rappel des souvenirs chez l'humain . Dans cette étude, 48 étudiants universitaires, hommes et femmes, en bonne santé, ont participé soit à un test de stress, soit à un groupe témoin. Les participants assignés aléatoirement au groupe stress ont eu la main immergée dans de l'eau glacée (le test de pression à froid socialement évalué, ou SECPT) pendant une durée maximale de trois minutes, sous surveillance et enregistrement vidéo. Les deux groupes (stress et témoin) ont ensuite dû mémoriser 32 mots. Vingt-quatre heures plus tard, les deux groupes ont été testés afin d'évaluer leur capacité à se souvenir de mots (rappel libre) et à en reconnaître un grand nombre parmi une liste plus longue (performance de reconnaissance). Les résultats ont révélé une nette altération des performances mnésiques dans le groupe soumis au stress, qui a rappelé 30 % de mots en moins que le groupe témoin. Les chercheurs suggèrent que le stress ressenti pendant l'apprentissage perturbe la concentration en détournant l'attention lors du processus d'encodage mnésique.
Cependant, les performances de la mémoire peuvent être améliorées lorsque le contenu est lié au contexte d'apprentissage, même en situation de stress. Une étude distincte menée par les psychologues cognitifs Schwabe et Wolf montre que lorsque les tests de rétention sont effectués dans un contexte similaire ou congruent avec la tâche d'apprentissage initiale (c'est-à-dire dans la même pièce), les troubles de la mémoire et les effets néfastes du stress sur l'apprentissage peuvent être atténués. Soixante-douze étudiants universitaires, hommes et femmes, en bonne santé, répartis aléatoirement entre le test de stress SECPT et un groupe témoin, ont été invités à mémoriser l'emplacement de 15 paires de cartes illustrées – une version informatisée du jeu de cartes « Concentration » ou « Mémoire ». La pièce où se déroulait l'expérience était imprégnée d'un parfum de vanille, l'odeur étant un puissant stimulus pour la mémoire. Les tests de rétention ont eu lieu le lendemain, soit dans la même pièce avec le même parfum de vanille, soit dans une autre pièce sans parfum. Les performances mnésiques des sujets ayant subi un stress lors de la tâche de localisation d'objets ont diminué significativement lorsqu'ils étaient testés dans une pièce inconnue sans parfum de vanille (contexte incongruent). En revanche, leurs performances mnésiques n'ont montré aucune altération lorsqu'ils étaient testés dans la pièce d'origine avec le parfum de vanille (contexte congruent). Tous les participants à l'expérience, stressés ou non, ont été plus rapides lorsque les contextes d'apprentissage et de récupération étaient similaires.
Ces recherches sur les effets du stress sur la mémoire pourraient avoir des implications pratiques pour l'éducation, le témoignage oculaire et la psychothérapie : les élèves pourraient obtenir de meilleurs résultats lors des tests effectués dans leur salle de classe habituelle plutôt que dans une salle d'examen, les témoins oculaires pourraient mieux se souvenir des détails sur les lieux d'un événement que dans une salle d'audience, et les personnes souffrant de stress post-traumatique pourraient aller mieux lorsqu'on les aide à situer leurs souvenirs d'un événement traumatique dans un contexte approprié.
Les expériences de vie stressantes peuvent être une cause de pertes de mémoire liées à l'âge. Les glucocorticoïdes libérés lors du stress endommagent les neurones de l' hippocampe . Ainsi, plus une personne est exposée au stress, plus elle est susceptible de souffrir de pertes de mémoire. Les neurones CA1 de l'hippocampe sont détruits par les glucocorticoïdes qui diminuent la libération de glucose et la recapture du glutamate . Ce taux élevé de glutamate extracellulaire permet au calcium de pénétrer dans les récepteurs NMDA, ce qui entraîne la mort des neurones. Les expériences de vie stressantes peuvent également provoquer un refoulement des souvenirs, une personne déplaçant un souvenir insupportable dans l'inconscient. Ce phénomène est directement lié à des événements traumatiques du passé, tels que des enlèvements, la captivité ou des abus sexuels durant l'enfance.
Plus l'exposition au stress est prolongée, plus son impact peut être important. Cependant, une exposition de courte durée au stress altère également la mémoire en perturbant le fonctionnement de l'hippocampe. Des recherches montrent que les sujets placés dans une situation stressante pendant une courte période présentent encore des taux de glucocorticoïdes sanguins fortement augmentés une fois l'exposition terminée. Lorsqu'on leur demande d'effectuer une tâche d'apprentissage après une exposition de courte durée, ils rencontrent souvent des difficultés. Le stress prénatal entrave également la capacité d'apprendre et de mémoriser en perturbant le développement de l'hippocampe et peut entraîner une potentialisation à long terme non établie chez les enfants de parents ayant subi un stress important. Bien que le stress soit appliqué avant la naissance, les enfants présentent des taux de glucocorticoïdes accrus lorsqu'ils sont soumis au stress plus tard dans leur vie. Une explication possible des performances mnésiques généralement inférieures des enfants issus de milieux socio-économiques défavorisés par rapport à leurs pairs issus de milieux plus aisés réside dans les effets du stress accumulé tout au long de leur vie. On pense également que les effets d'un faible revenu sur le développement de l'hippocampe sont médiés par des réponses au stress chronique, ce qui pourrait expliquer pourquoi les enfants issus de milieux à faibles et à hauts revenus diffèrent en termes de performances mnésiques.
Dormir
La formation des souvenirs se déroule en trois étapes, un processus que le sommeil peut favoriser . Ces trois étapes sont les suivantes :
- L'acquisition est le processus de stockage et de récupération de nouvelles informations en mémoire.
- Consolidation
- Rappel
Le sommeil influence la consolidation de la mémoire. Pendant le sommeil, les connexions neuronales du cerveau se renforcent, ce qui améliore sa capacité à stabiliser et à retenir les souvenirs. Plusieurs études ont démontré que le sommeil améliore la mémorisation, car les souvenirs sont renforcés par une consolidation active. La consolidation systémique a lieu pendant le sommeil lent profond (SLP). Ce processus implique que les souvenirs sont réactivés pendant le sommeil, mais que ce processus ne renforce pas tous les souvenirs. Il implique également que des modifications qualitatives sont apportées aux souvenirs lors de leur transfert vers la mémoire à long terme pendant le sommeil. Pendant le sommeil, l'hippocampe rejoue les événements de la journée pour le néocortex. Le néocortex examine et traite ensuite les souvenirs, ce qui les transfère dans la mémoire à long terme. Un manque de sommeil rend l'apprentissage plus difficile car ces connexions neuronales sont moins fortes, ce qui entraîne un taux de rétention des souvenirs plus faible. La privation de sommeil rend la concentration plus difficile, ce qui nuit à l'apprentissage. De plus, certaines études ont montré que le manque de sommeil peut entraîner la formation de faux souvenirs , car ces derniers ne sont pas correctement transférés vers la mémoire à long terme. L'une des principales fonctions du sommeil serait d'améliorer la consolidation des informations, car plusieurs études ont démontré que la mémoire dépend d'un sommeil suffisant entre l'apprentissage et l'évaluation. Par ailleurs, des données issues d'études de neuro-imagerie ont révélé des schémas d'activation cérébrale pendant le sommeil qui reflètent ceux enregistrés lors de l'apprentissage des tâches la veille, suggérant que les nouveaux souvenirs pourraient être consolidés par cette répétition.
Construction pour manipulation générale
Bien que l'on imagine souvent la mémoire comme un simple enregistreur, ce n'est pas le cas. Les mécanismes moléculaires sous-jacents à l'induction et au maintien de la mémoire sont très dynamiques et comprennent des phases distinctes s'étalant sur une période allant de quelques secondes à toute une vie. En réalité, la recherche a révélé que nos souvenirs sont construits : « les hypothèses actuelles suggèrent que les processus constructifs permettent aux individus de simuler et d'imaginer des épisodes futurs, des événements et des scénarios. Puisque le futur n'est pas une répétition exacte du passé, la simulation d'épisodes futurs requiert un système complexe capable de puiser dans le passé de manière à extraire et recombiner avec souplesse des éléments d'expériences antérieures – un système constructif plutôt que reproductif. » On peut construire ses souvenirs lors de leur encodage et/ou de leur rappel. À titre d'exemple, prenons l'étude classique d' Elizabeth Loftus et John Palmer (1974) dans laquelle des participants devaient visionner un film montrant un accident de la route, puis étaient interrogés sur ce qu'ils avaient vu. Les chercheurs ont constaté que les personnes interrogées sur la vitesse des voitures lors de leur collision ont donné des estimations plus élevées que celles interrogées sur la vitesse des voitures lors de leur impact . De plus, une semaine plus tard, lorsqu'on leur a demandé s'ils avaient vu des bris de verre dans le film, celles qui avaient entendu la question avec « bris de verre » étaient deux fois plus susceptibles de répondre par l'affirmative que celles qui avaient entendu la question avec « impact » (aucun bris de verre n'étant montré dans le film). Ainsi, la formulation des questions a influencé la mémoire des spectateurs. Plus important encore, cette formulation a conduit à la construction de souvenirs différents : celles qui avaient entendu la question avec « bris de verre » se souvenaient d' un accident de voiture plus grave que celui qu'elles avaient réellement vu. Les résultats de cette expérience ont été reproduits dans le monde entier, et les chercheurs ont systématiquement démontré que la désinformation induit une tendance à la déformation des souvenirs, un phénomène connu sous le nom d' effet de désinformation .
Des recherches ont révélé que demander à des individus d'imaginer à plusieurs reprises des actions qu'ils n'ont jamais accomplies ou des événements qu'ils n'ont jamais vécus pouvait engendrer de faux souvenirs. Par exemple, Goff et Roediger (1998) ont demandé à des participants d'imaginer avoir accompli une action (par exemple, casser un cure-dent), puis leur ont demandé ultérieurement s'ils l'avaient réellement faite. Les résultats ont montré que les participants qui avaient imaginé à plusieurs reprises accomplir cette action étaient plus susceptibles de croire l'avoir réellement accomplie lors de la première séance de l'expérience. De même, Garry et ses collègues (1996) ont demandé à des étudiants d'indiquer leur degré de certitude quant à la survenue de certains événements durant leur enfance (par exemple, casser une vitre avec la main), puis, deux semaines plus tard, leur ont demandé d'imaginer quatre de ces événements. Les chercheurs ont constaté qu'un quart des étudiants invités à imaginer ces quatre événements ont déclaré les avoir réellement vécus durant leur enfance. Autrement dit, lorsqu'on leur demandait d'imaginer les événements, ils étaient plus confiants de les avoir vécus.
Des recherches publiées en 2013 ont révélé qu'il est possible de stimuler artificiellement des souvenirs antérieurs et d'implanter artificiellement de faux souvenirs chez la souris. Grâce à l'optogénétique , une équipe de scientifiques du RIKEN et du MIT a amené ces souris à associer, à tort, un environnement rassurant à une expérience désagréable vécue dans un autre contexte. Certains scientifiques estiment que cette étude pourrait avoir des implications pour l'étude de la formation de faux souvenirs chez l'humain, ainsi que pour le traitement du syndrome de stress post-traumatique ( SSPT) et de la schizophrénie .
La reconsolidation de la mémoire se produit lorsque des souvenirs précédemment consolidés sont rappelés ou récupérés de la mémoire à long terme vers la conscience active. Au cours de ce processus, les souvenirs peuvent être renforcés et enrichis, mais il existe également un risque de manipulation. Nous avons tendance à considérer nos souvenirs comme stables et constants lorsqu'ils sont stockés dans la mémoire à long terme, mais ce n'est pas le cas. De nombreuses études ont montré que la consolidation des souvenirs n'est pas un événement unique, mais qu'ils subissent un processus répété, appelé reconsolidation. C'est lorsque le souvenir est rappelé ou récupéré et replacé dans la mémoire de travail. Le souvenir est alors vulnérable à la manipulation par des sources externes et à l'effet de désinformation, qui peut être dû à une mauvaise attribution de la source de l'information incohérente, avec ou sans trace mnésique originale intacte. Une chose est sûre : la mémoire est malléable.
Ces nouvelles recherches sur le concept de reconsolidation ont ouvert la voie à des méthodes pour aider les personnes souffrant de souvenirs désagréables ou ayant des difficultés de mémorisation. Par exemple, si vous avez vécu une expérience traumatisante et que vous vous remémorez ce souvenir dans un environnement moins stimulant, il sera affaibli lors de sa prochaine récupération. « Certaines études suggèrent que les souvenirs sur-entraînés ou fortement renforcés ne subissent pas de reconsolidation s'ils sont réactivés dans les premiers jours suivant l'entraînement, mais deviennent sensibles aux interférences de reconsolidation au fil du temps. » Cela ne signifie toutefois pas que tous les souvenirs sont susceptibles de reconsolidation. Il existe des preuves suggérant que les souvenirs ayant fait l'objet d'un entraînement intensif, qu'il soit intentionnel ou non, sont moins susceptibles de se reconsolider. Des tests supplémentaires menés sur des rats et dans des labyrinthes ont montré que les souvenirs réactivés étaient plus sensibles à la manipulation, positive comme négative, que les souvenirs nouvellement formés. On ignore encore s’il s’agit de nouveaux souvenirs formés, et d’une incapacité à récupérer le souvenir approprié à la situation, ou s’il s’agit d’un souvenir reconsolidé. L’étude de la reconsolidation étant un concept relativement récent, sa validité scientifique fait encore débat.
Amélioration
Une étude de l'UCLA, publiée dans le numéro de juin 2008 de l' American Journal of Geriatric Psychiatry, a démontré que de simples changements de mode de vie, comme l'intégration d'exercices de mémoire, d' une alimentation saine , d'une activité physique régulière et la réduction du stress, permettent d'améliorer les fonctions cognitives et l'efficacité cérébrale . Cette étude a porté sur 17 sujets (âge moyen : 53 ans) présentant des performances mnésiques normales. Huit d'entre eux ont suivi un régime alimentaire favorisant la santé cérébrale, incluant des exercices de relaxation, d'activité physique et de stimulation mentale (énigmes et techniques d'entraînement de la mémoire verbale ). Après 14 jours, leur fluidité verbale (et non leur mémoire) s'est améliorée par rapport à leurs performances initiales. Aucun suivi à long terme n'ayant été réalisé, il est impossible de déterminer si cette intervention a des effets durables sur la mémoire.
L’exercice physique, même d’intensité légère, améliore significativement la mémoire à tous les âges, les bénéfices les plus importants étant observés chez les enfants et les adolescents. Même un exercice d’intensité faible à modérée et des interventions plus courtes (1 à 3 mois) peuvent produire des améliorations cognitives significatives.
Il existe un ensemble assez vague de principes et de techniques mnémotechniques qui peuvent être utilisés pour améliorer considérablement la mémoire, connu sous le nom d' art de la mémoire .
Le Centre international de la longévité a publié en 2001 un rapport qui comprend, aux pages 14 à 16, des recommandations pour préserver les fonctions cognitives jusqu'à un âge avancé. Voici quelques-unes de ces recommandations :
- se maintenir intellectuellement actif par l'apprentissage, la formation ou la lecture
- rester physiquement actif afin de favoriser la circulation sanguine vers le cerveau
- socialiser
- pour réduire le stress
- pour maintenir un horaire de sommeil régulier
- pour éviter la dépression ou l'instabilité émotionnelle
- observer une bonne nutrition.
La mémorisation est une méthode d'apprentissage qui permet de se souvenir d'informations mot pour mot. L'apprentissage par cœur est la méthode la plus couramment utilisée. Les méthodes de mémorisation ont fait l'objet de nombreuses discussions au fil des ans, certains auteurs, comme Cosmos Rossellius, ayant utilisé des alphabets visuels . L' effet d'espacement montre qu'une personne est plus susceptible de se souvenir d'une liste d'éléments lorsque la répétition est espacée dans le temps. À l'inverse , le bachotage consiste en une mémorisation intensive sur une courte période. L'effet d'espacement est exploité pour améliorer la mémoire grâce à la méthode des flashcards par répétition espacée . L' effet Zeigarnik est également pertinent : il stipule que l'on se souvient mieux des tâches inachevées ou interrompues que des tâches terminées. La méthode des lieux utilise la mémoire spatiale pour mémoriser des informations non spatiales.
différences entre les sexes
Les recherches sur les différences de mémoire épisodique entre les sexes ont donné des résultats mitigés. Une méta-analyse récente a révélé un léger avantage global chez les femmes, avec des variations selon les tâches. Les femmes ont obtenu de meilleurs résultats que les hommes lors des tâches de rappel indicé et de rappel libre, tandis que les hommes ont montré un avantage dans les tâches d'empan complexe. Aucune différence entre les sexes n'a été observée lors des tâches de rappel sériel ou d'empan simple. Des facteurs tels que le sens du rappel, le type de stimulus, le format de présentation, le format de réponse et l'âge expliquent la variance des résultats. Il est important de noter qu'aucun biais de publication n'a été détecté, bien que la taille des effets ait varié selon la source de l'échantillon et la méthodologie de l'étude. Des études de neuro-imagerie utilisant l'estimation de la vraisemblance d'activation (ALE) ont indiqué une activité plus importante chez les hommes que chez les femmes dans le cortex préfrontal latéral, les régions visuelles, le cortex parahippocampique et le cervelet lors de la récupération de la mémoire à long terme. Ces résultats suggèrent des différences significatives entre les sexes, tant au niveau du comportement que du fonctionnement cérébral, soulignant la nécessité d'une interprétation prudente et de recherches contrôlées supplémentaires.
Chez les plantes
Les plantes étant dépourvues d'organe spécialisé dans la mémorisation, la mémoire végétale a fait l'objet de controverses ces dernières années. De récentes avancées dans ce domaine ont mis en évidence la présence de neurotransmetteurs chez les plantes, renforçant ainsi l'hypothèse de leur capacité de mémorisation. Il a également été démontré que les potentiels d'action , une réponse physiologique caractéristique des neurones , influencent les plantes, notamment la réponse aux blessures et la photosynthèse . Outre ces caractéristiques communes aux systèmes de mémoire des plantes et des animaux, on a également observé que les plantes encodent, stockent et récupèrent des souvenirs à court terme élémentaires.
L'une des plantes les plus étudiées pour ses propriétés de mémoire rudimentaire est la dionée attrape-mouche . Originaire des zones humides subtropicales de l' est des États-Unis , cette plante a développé la capacité de se nourrir de viande, probablement en raison de la pauvreté du sol en azote. Elle y parvient grâce à deux lobes foliaires qui se referment brusquement au contact d'une proie potentielle. Sur chaque lobe, trois poils sensitifs sont prêts à être stimulés. Afin d'optimiser le rapport bénéfice/coût, la plante possède une forme rudimentaire de mémoire : deux poils sensitifs doivent être stimulés en moins de trente secondes pour que le piège se referme. Ce système garantit que le piège ne se referme que lorsqu'une proie potentielle est à portée.
L'intervalle de temps entre les stimulations des poils sensitifs suggère que la plante peut se souvenir d'un stimulus initial suffisamment longtemps pour qu'un second stimulus déclenche la fermeture du piège. Cette mémoire n'est pas encodée dans un cerveau, car les plantes sont dépourvues de cet organe spécialisé. L'information est plutôt stockée sous forme de niveaux de calcium cytoplasmique . Le premier stimulus provoque un influx de calcium cytoplasmique inférieur au seuil. Ce stimulus initial n'est pas suffisant pour activer la fermeture du piège ; un stimulus ultérieur permet donc un second influx de calcium. Cette seconde augmentation de calcium se superpose à la première, créant un potentiel d'action qui dépasse le seuil, entraînant la fermeture du piège. Afin de prouver qu'un seuil électrique doit être atteint pour stimuler la fermeture du piège, des chercheurs ont excité un seul poil sensitif par un stimulus mécanique constant à l'aide d'électrodes Ag/AgCl. Le piège s'est fermé après seulement quelques secondes. Cette expérience a démontré que le seuil électrique, et non nécessairement le nombre de stimulations des poils sensitifs, était le facteur déterminant de la mémoire de la dionée attrape-mouche.
Il a été démontré que la fermeture du piège peut être bloquée à l'aide de découpleurs et d'inhibiteurs des canaux voltage-dépendants . Après la fermeture du piège, ces signaux électriques stimulent la production glandulaire d' acide jasmonique et d'hydrolases , permettant la digestion des proies.
De nombreuses autres plantes présentent une capacité de mémorisation, notamment Mimosa pudica . Un dispositif expérimental a été conçu pour laisser tomber des plants de mimosa en pot à plusieurs reprises, d'une même hauteur et à la même vitesse. On a observé que la réaction défensive des plantes, consistant à enrouler leurs feuilles, diminuait au fil des soixante répétitions de l'expérience. Pour confirmer qu'il s'agissait d'un mécanisme de mémorisation et non d'un phénomène d' épuisement , certains plants ont été secoués après l'expérience et ont présenté une réaction défensive normale, à savoir l'enroulement des feuilles. Cette expérience a démontré l'existence d'une mémoire à long terme chez les plantes, car elle a été répétée un mois plus tard et les plants sont restés insensibles à la chute.