Le séquençage de Sanger est une méthode de séquençage de l'ADN qui implique l'électrophorèse et est basée sur l'incorporation aléatoire de didésoxynucléotides de terminaison de chaîne par l'ADN polymérase pendant la réplication de l'ADN in vitro . Après avoir été développé par Frederick Sanger et ses collègues en 1977, il est devenu la méthode de séquençage la plus utilisée pendant environ 40 ans. Un instrument automatisé utilisant l'électrophorèse sur gel en plaque et des marqueurs fluorescents a été commercialisé pour la première fois par Applied Biosystems en mars 1987. Plus tard, les gels en plaque automatisés ont été remplacés par l'électrophorèse sur réseau capillaire automatisée. Plus récemment, le séquençage de Sanger à volume plus élevé a été remplacé par des méthodes de séquençage de nouvelle génération , en particulier pour les analyses génomiques automatisées à grande échelle . Cependant, la méthode Sanger reste largement utilisée pour les projets à plus petite échelle et pour la validation des résultats de séquençage en profondeur. Il présente toujours l'avantage par rapport aux technologies de séquençage à lecture courte (comme Illumina) dans la mesure où il peut produire des lectures de séquences d'ADN de > 500 nucléotides et maintient un taux d'erreur très faible avec une précision d'environ 99,99 %. Le séquençage Sanger est toujours activement utilisé dans les efforts des initiatives de santé publique telles que le séquençage de la protéine de pointe du SRAS-CoV-2 ainsi que pour la surveillance des épidémies de norovirus par le biais du réseau de surveillance CaliciNet du Center for Disease Control and Prevention (CDC).
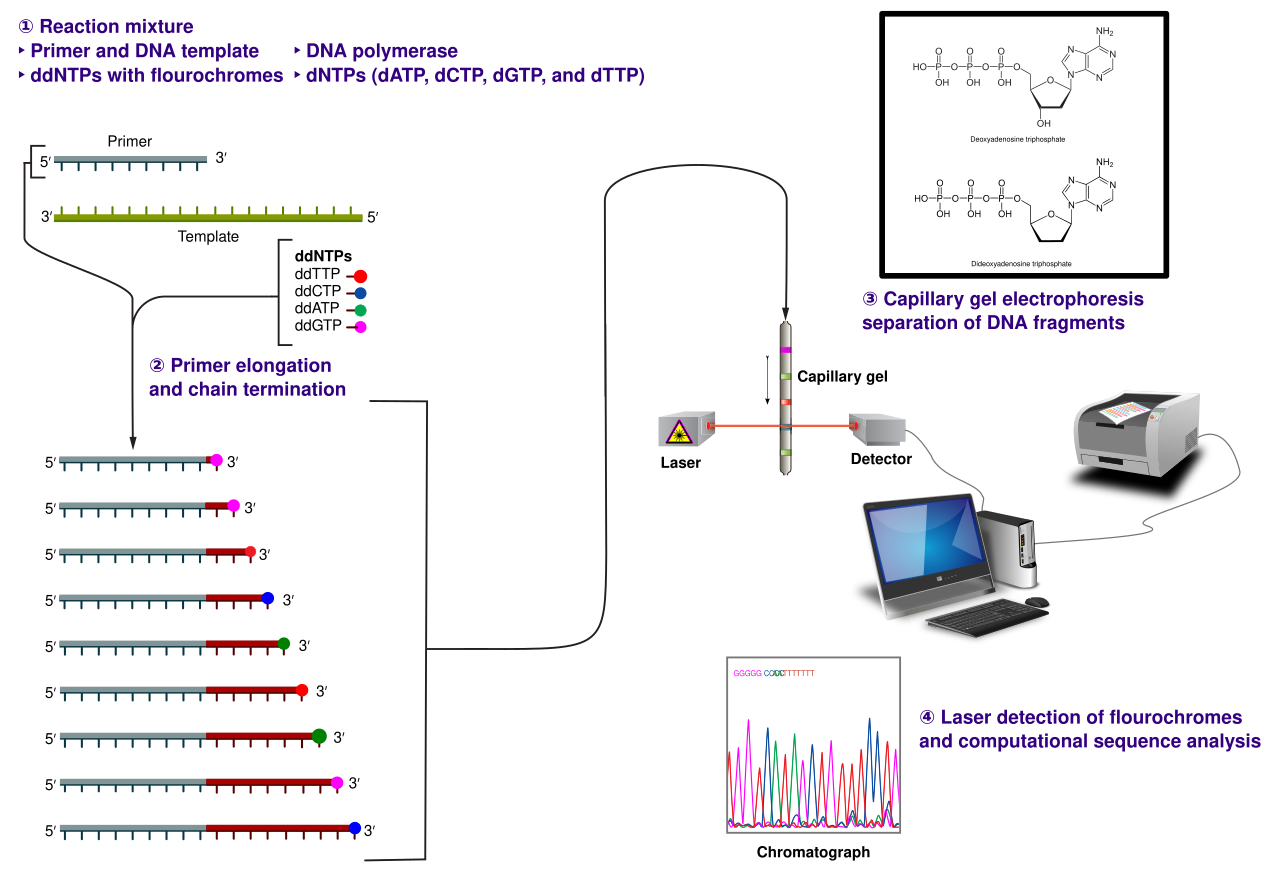
Méthode
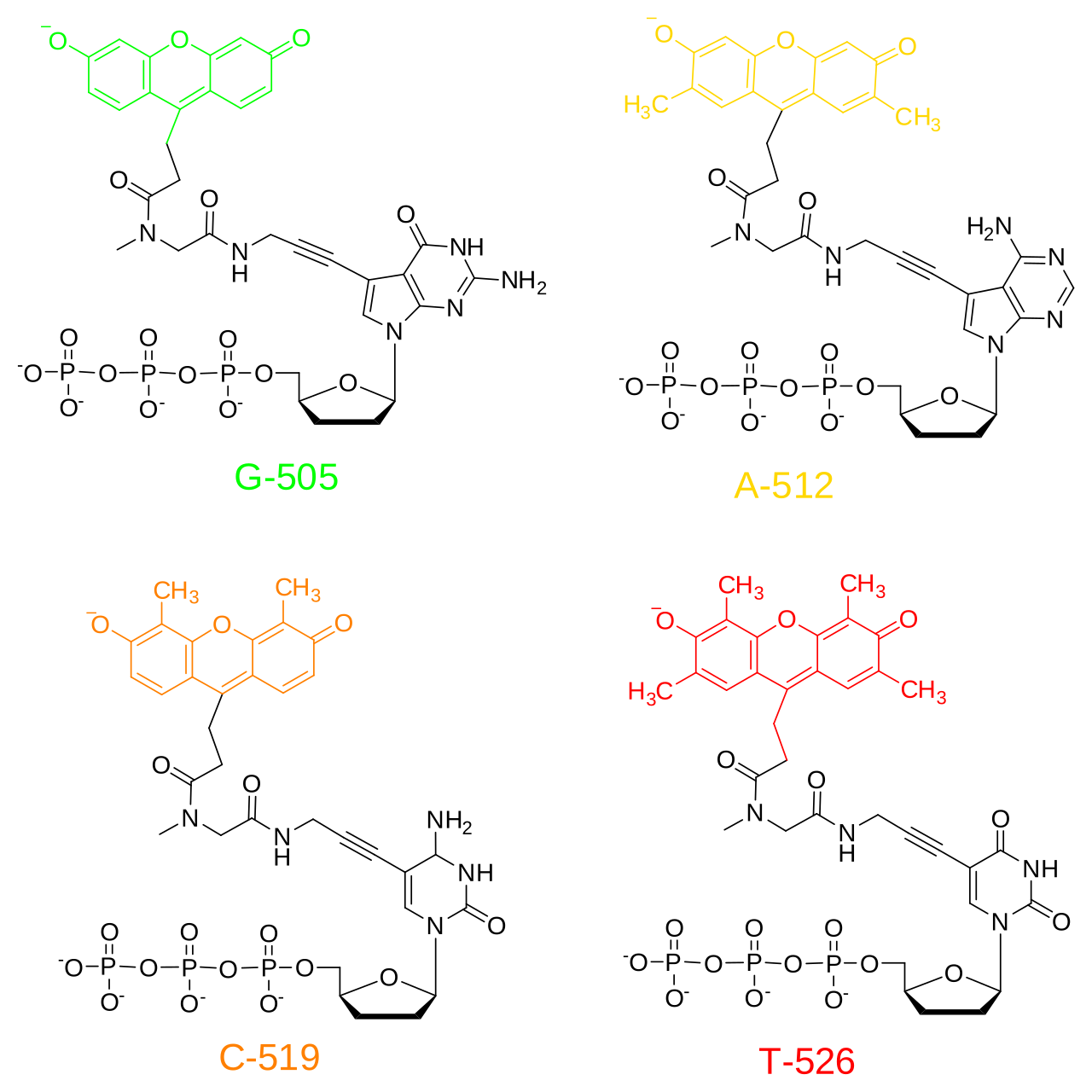
La méthode classique de terminaison de chaîne nécessite un modèle d'ADN monocaténaire, une amorce d'ADN , une ADN polymérase , des désoxynucléotides triphosphates normaux ( dNTP ) et des di-désoxynucléotides triphosphates modifiés ( ddNTP ), ces derniers interrompant l'élongation du brin d'ADN. Ces nucléotides de terminaison de chaîne sont dépourvus du groupe 3'- OH nécessaire à la formation d'une liaison phosphodiester entre deux nucléotides, ce qui fait que l'ADN polymérase cesse l'extension de l'ADN lorsqu'un ddNTP modifié est incorporé. Les ddNTP peuvent être marqués par radioactivité ou par fluorescence pour être détectés dans des machines de séquençage automatisées.
L'échantillon d'ADN est divisé en quatre réactions de séquençage distinctes, contenant les quatre désoxynucléotides standards (dATP, dGTP, dCTP et dTTP) et l'ADN polymérase. À chaque réaction est ajouté un seul des quatre didésoxynucléotides (ddATP, ddGTP, ddCTP ou ddTTP), tandis que les autres nucléotides ajoutés sont des nucléotides ordinaires. La concentration en désoxynucléotides doit être environ 100 fois supérieure à celle du didésoxynucléotide correspondant (par exemple 0,5 mM dTTP : 0,005 mM ddTTP) pour permettre la production de suffisamment de fragments tout en transcrivant la séquence complète (mais la concentration en ddNTP dépend également de la longueur de séquence souhaitée). Pour mettre les choses dans un ordre plus raisonnable, quatre réactions distinctes sont nécessaires dans ce processus pour tester les quatre ddNTP. Après des cycles d'extension de l'ADN modèle à partir de l'amorce liée, les fragments d'ADN résultants sont dénaturés par la chaleur et séparés par taille à l'aide d'une électrophorèse sur gel . Dans la publication originale de 1977, la formation de boucles de paires de bases d'ADN simple brin était une cause de sérieuses difficultés pour résoudre les bandes à certains endroits. Cela est souvent réalisé en utilisant un gel de polyacrylamide -urée dénaturant avec chacune des quatre réactions exécutées dans l'une des quatre voies individuelles (voies A, T, G, C). Les bandes d'ADN peuvent ensuite être visualisées par autoradiographie ou lumière UV, et la séquence d'ADN peut être lue directement sur le film radiographique ou l'image du gel.
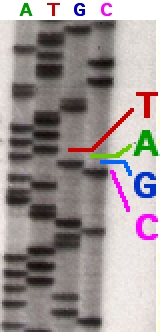
Dans l'image de droite, le film radiographique a été exposé au gel et les bandes sombres correspondent à des fragments d'ADN de différentes longueurs. Une bande sombre dans une voie indique un fragment d'ADN qui est le résultat de la terminaison de chaîne après l'incorporation d'un didésoxynucléotide (ddATP, ddGTP, ddCTP ou ddTTP). Les positions relatives des différentes bandes parmi les quatre voies, de bas en haut, sont ensuite utilisées pour lire la séquence d'ADN.

Les variantes techniques du séquençage de terminaison de chaîne comprennent le marquage avec des nucléotides contenant du phosphore radioactif pour le radiomarquage , ou l'utilisation d'une amorce marquée à l'extrémité 5' avec un colorant fluorescent . Le séquençage par amorce de colorant facilite la lecture dans un système optique pour une analyse et une automatisation plus rapides et plus économiques. Le développement ultérieur par Leroy Hood et ses collègues de ddNTP et d'amorces marqués par fluorescence a ouvert la voie au séquençage automatisé de l'ADN à haut débit.
Les méthodes de terminaison de chaîne ont grandement simplifié le séquençage de l'ADN. Par exemple, des kits basés sur la terminaison de chaîne sont disponibles dans le commerce et contiennent les réactifs nécessaires au séquençage, pré-aliquotés et prêts à l'emploi. Les limitations incluent la liaison non spécifique de l'amorce à l'ADN, affectant la lecture précise de la séquence d'ADN, et les structures secondaires de l'ADN affectant la fidélité de la séquence.
Séquençage par terminateur de colorant
Le séquençage par terminateur de colorant utilise le marquage des ddNTP terminateurs de chaîne, ce qui permet le séquençage en une seule réaction plutôt qu'en quatre réactions comme dans la méthode des amorces marquées. Dans le séquençage par terminateur de colorant, chacun des quatre terminateurs de chaîne didésoxynucléotidiques est marqué avec des colorants fluorescents, chacun d'eux émettant de la lumière à des longueurs d'onde différentes .
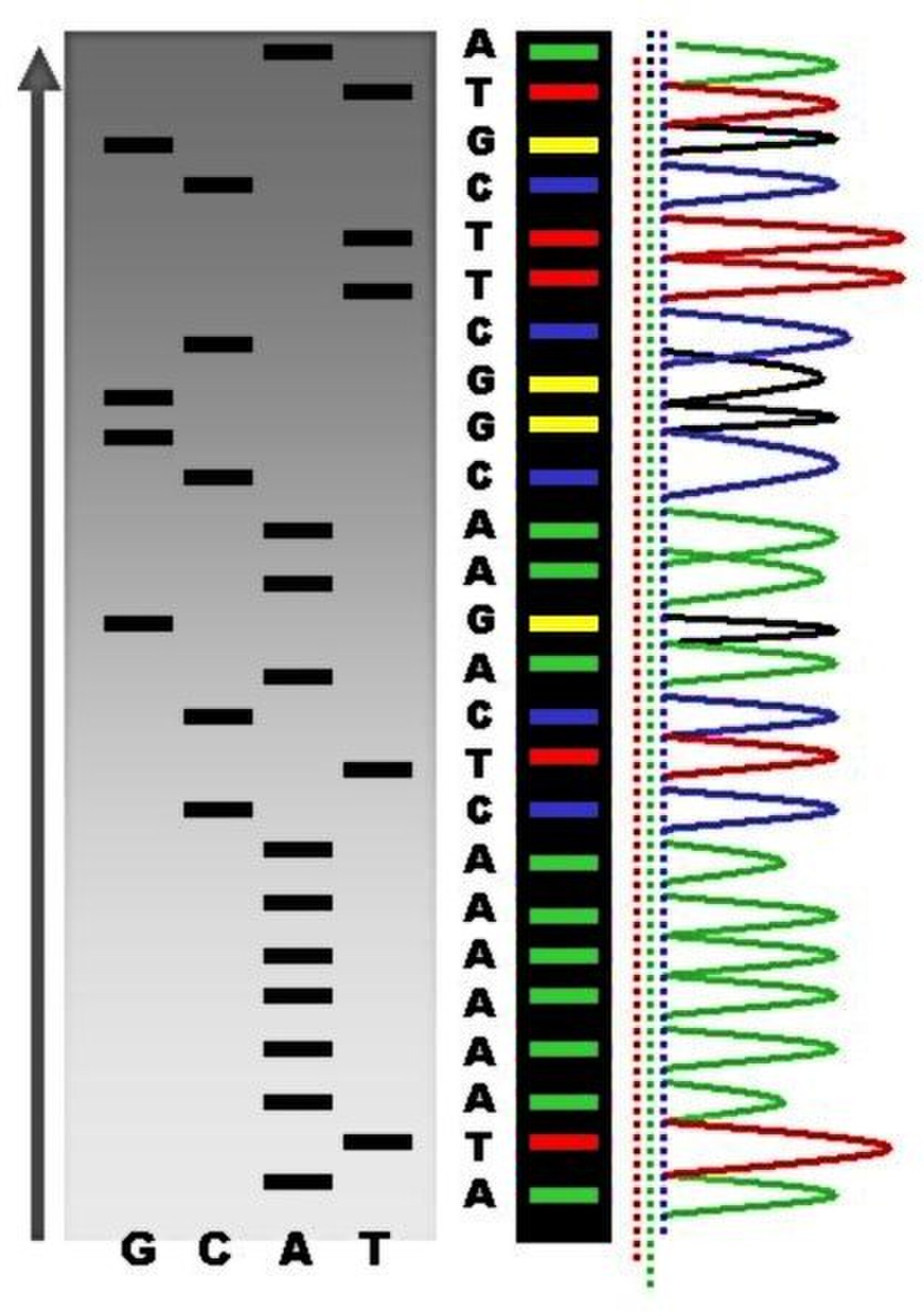
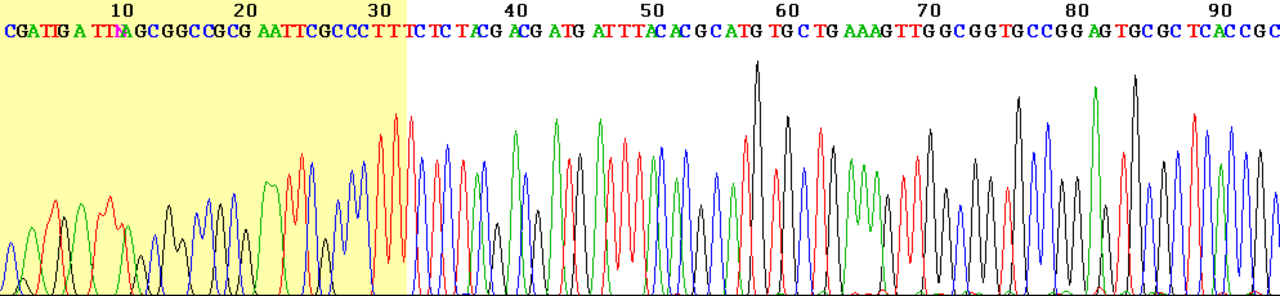
En raison de sa plus grande rapidité et de sa plus grande efficacité, le séquençage par terminateur de colorant est aujourd'hui le pilier du séquençage automatisé. Ses limites incluent les effets du colorant dus aux différences d'incorporation des terminateurs de chaîne marqués par colorant dans le fragment d'ADN, ce qui entraîne des hauteurs et des formes de pics inégales dans l' électrophorégramme de trace de séquence d'ADN électronique (un type de chromatogramme ) après électrophorèse capillaire (voir la figure de gauche).
Ce problème a été résolu grâce à l'utilisation de systèmes d'enzymes polymérases d'ADN modifiées et de colorants qui minimisent la variabilité d'incorporation, ainsi que de méthodes permettant d'éliminer les « taches de colorant ». La méthode de séquençage par terminateur de colorant, ainsi que les analyseurs de séquences d'ADN automatisés à haut débit, ont été utilisés pour la grande majorité des projets de séquençage jusqu'à l'introduction du séquençage de nouvelle génération .
Automatisation et préparation d'échantillons
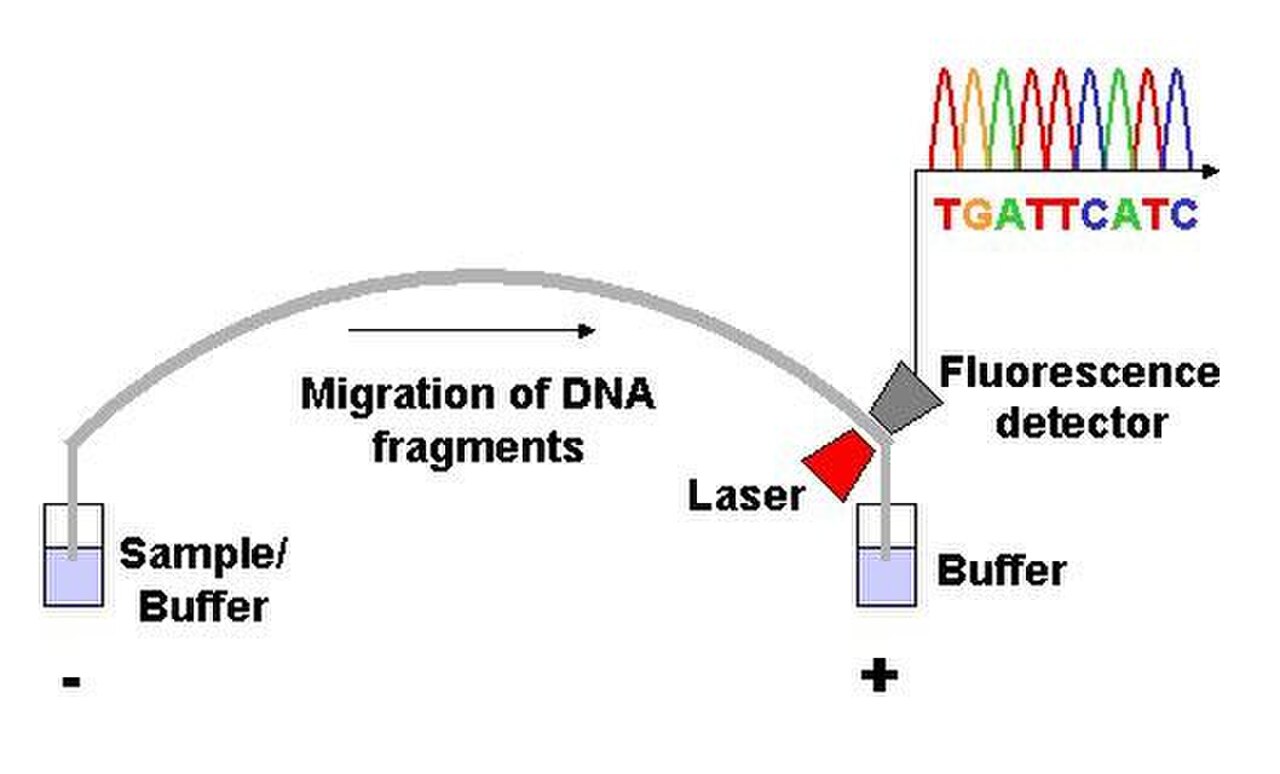
Les instruments de séquençage d'ADN automatisés ( séquenceurs d'ADN ) peuvent séquencer jusqu'à 384 échantillons d'ADN en un seul lot. Les exécutions par lots peuvent avoir lieu jusqu'à 24 fois par jour. Les séquenceurs d'ADN séparent les brins par taille (ou longueur) à l'aide d' une électrophorèse capillaire , ils détectent et enregistrent la fluorescence des colorants et génèrent des données sous forme de chromatogrammes de traces de pics fluorescents . Les réactions de séquençage ( thermocyclage et marquage), le nettoyage et la remise en suspension des échantillons dans une solution tampon sont effectués séparément, avant le chargement des échantillons sur le séquenceur. Un certain nombre de logiciels commerciaux et non commerciaux peuvent éliminer automatiquement les traces d'ADN de mauvaise qualité. Ces programmes évaluent la qualité de chaque pic et suppriment les pics de base de mauvaise qualité (qui sont généralement situés aux extrémités de la séquence). La précision de ces algorithmes est inférieure à l'examen visuel par un opérateur humain, mais est adéquate pour le traitement automatisé de grands ensembles de données de séquence.
Applications du séquençage avec terminaison par colorant
Le domaine de la santé publique joue de nombreux rôles pour soutenir le diagnostic des patients ainsi que la surveillance environnementale des substances toxiques potentielles et des agents pathogènes biologiques en circulation. Les laboratoires de santé publique (PHL) et d'autres laboratoires du monde entier ont joué un rôle central en fournissant des données de séquençage rapide pour la surveillance du virus SARS-CoV-2 , agent causal de la COVID-19, pendant la pandémie qui a été déclarée urgence de santé publique le 30 janvier 2020. Les laboratoires ont été chargés de la mise en œuvre rapide de méthodes de séquençage et ont été invités à fournir des données précises pour aider aux modèles de prise de décision pour l'élaboration de politiques visant à atténuer la propagation du virus. De nombreux laboratoires ont eu recours à des méthodologies de séquençage de nouvelle génération tandis que d'autres ont soutenu les efforts de séquençage Sanger. Les efforts de séquençage du SARS-CoV-2 sont nombreux, alors que la plupart des laboratoires ont mis en œuvre le séquençage du génome entier du virus, d'autres ont choisi de séquencer des gènes très spécifiques du virus tels que le gène S, codant les informations nécessaires à la production de la protéine de pointe. Le taux de mutation élevé du SARS-CoV-2 entraîne des différences génétiques au sein du gène S et ces différences ont joué un rôle dans l'infectiosité du virus. Le séquençage Sanger du gène S offre une méthode rapide, précise et plus abordable pour récupérer le code génétique. Les laboratoires des pays à faible revenu n'ont peut-être pas les capacités de mettre en œuvre des applications coûteuses telles que le séquençage de nouvelle génération, de sorte que les méthodes Sanger peuvent prévaloir pour soutenir la génération de données de séquençage pour la surveillance des variants.
Le séquençage Sanger est également la « référence absolue » des méthodes de surveillance des norovirus pour le réseau CaliciNet du Centre de contrôle et de prévention des maladies (CDC). CalciNet est un réseau de surveillance des épidémies qui a été créé en mars 2009. L'objectif du réseau est de collecter des données de séquençage des norovirus en circulation aux États-Unis et d'activer des mesures en aval pour déterminer la source de l'infection afin d'atténuer la propagation du virus. Le réseau CalciNet a identifié de nombreuses infections comme des maladies d'origine alimentaire. Ces données peuvent ensuite être publiées et utilisées pour élaborer des recommandations pour les actions futures visant à prévenir la contamination des aliments. Les méthodes employées pour la détection des norovirus impliquent l'amplification ciblée de zones spécifiques du génome. Les amplicons sont ensuite séquencés à l'aide du séquençage Sanger à terminaison de colorant et les chromatogrammes et séquences générés sont analysés avec un progiciel développé par BioNumerics . Les séquences sont suivies et la parenté des souches est étudiée pour en déduire la pertinence épidémiologique.
Défis
Les défis courants du séquençage de l'ADN avec la méthode Sanger incluent la mauvaise qualité des 15 à 40 premières bases de la séquence en raison de la liaison de l'amorce et la détérioration de la qualité des traces de séquençage après 700 à 900 bases. Les logiciels d'appel de bases tels que Phred fournissent généralement une estimation de la qualité pour aider à la suppression des régions de séquences de faible qualité.
Dans les cas où les fragments d'ADN sont clonés avant le séquençage, la séquence résultante peut contenir des parties du vecteur de clonage . En revanche, les technologies de clonage basées sur la PCR et de séquençage de nouvelle génération basées sur le pyroséquençage évitent souvent l'utilisation de vecteurs de clonage. Récemment, des méthodes de séquençage Sanger en une étape (amplification et séquençage combinés) telles qu'Ampliseq et SeqSharp ont été développées qui permettent un séquençage rapide des gènes cibles sans clonage ni amplification préalable.
Les méthodes actuelles ne permettent de séquencer directement que des fragments d'ADN relativement courts (300 à 1000 nucléotides de long) en une seule réaction. Le principal obstacle au séquençage de fragments d'ADN dépassant cette limite de taille est le manque de puissance de séparation pour distinguer les grands fragments d'ADN dont la longueur ne diffère que d'un nucléotide.
Séquençage Sanger microfluidique
Le séquençage microfluidique Sanger est une application de séquençage d'ADN en laboratoire sur puce , dans laquelle les étapes de séquençage Sanger (cyclage thermique, purification d'échantillons et électrophorèse capillaire) sont intégrées sur une puce à l'échelle d'une tranche de wafer en utilisant des volumes d'échantillons à l'échelle du nanolitre. Cette technologie génère des lectures de séquence longues et précises, tout en évitant de nombreuses lacunes importantes de la méthode Sanger conventionnelle (par exemple, une consommation élevée de réactifs coûteux, la dépendance à un équipement coûteux, des manipulations nécessitant beaucoup de personnel, etc.) en intégrant et en automatisant les étapes de séquençage Sanger.
Dans sa conception moderne, le séquençage du génome à haut débit implique la fragmentation du génome en petits morceaux monocaténaires, suivie de l'amplification des fragments par réaction en chaîne par polymérase (PCR). En adoptant la méthode Sanger, chaque fragment d'ADN est terminé de manière irréversible par l'incorporation d'un nucléotide de terminaison de chaîne didésoxy marqué par fluorescence, produisant ainsi une « échelle » d'ADN de fragments qui diffèrent chacun en longueur d'une base et portent un marqueur fluorescent spécifique à la base terminale. Les échelles de bases amplifiées sont ensuite séparées par électrophorèse capillaire (CAE) avec détection automatisée in situ de la « ligne d'arrivée » des fragments d'ADN simple brin marqués par fluorescence, ce qui fournit une séquence ordonnée des fragments. Ces lectures de séquences sont ensuite assemblées par ordinateur en séquences superposées ou contiguës (appelées « contigs ») qui ressemblent à la séquence génomique complète une fois entièrement assemblées.
Les méthodes Sanger atteignent des longueurs de lecture maximales d'environ 800 pb (généralement 500 à 600 pb avec de l'ADN non enrichi). Les longueurs de lecture plus longues dans les méthodes Sanger présentent des avantages significatifs par rapport aux autres méthodes de séquençage, notamment en termes de séquençage de régions répétitives du génome. Le défi des données de séquences de lecture courtes est particulièrement important dans le séquençage de nouveaux génomes (de novo) et dans le séquençage de segments de génomes fortement réorganisés, généralement ceux observés dans les génomes cancéreux ou dans les régions de chromosomes qui présentent une variation structurelle.
Applications des technologies de séquençage microfluidique
D'autres applications utiles du séquençage de l'ADN comprennent la détection du polymorphisme d'un seul nucléotide (SNP), l'analyse hétéroduplex du polymorphisme de conformation d'un seul brin (SSCP) et l'analyse des répétitions courtes en tandem (STR). La résolution des fragments d'ADN en fonction des différences de taille et/ou de conformation est l'étape la plus critique dans l'étude de ces caractéristiques du génome.
Conception de l'appareil
La puce de séquençage est constituée de quatre couches, constituées de trois plaquettes de verre de 100 mm de diamètre (sur lesquelles les éléments du dispositif sont microfabriqués) et d'une membrane en polydiméthylsiloxane (PDMS). Les chambres de réaction et les canaux d'électrophorèse capillaire sont gravés entre les deux plaquettes de verre supérieures, qui sont liées thermiquement. Les interconnexions de canaux tridimensionnelles et les microvannes sont formées par le PDMS et la plaquette de verre du collecteur inférieur.
Le dispositif se compose de trois unités fonctionnelles, chacune correspondant aux étapes du séquençage Sanger. L'unité de cyclage thermique (TC) est une chambre de réaction de 250 nanolitres avec détecteur de température résistif intégré, microvannes et un élément chauffant de surface. Le mouvement du réactif entre la couche supérieure entièrement en verre et la couche inférieure en verre-PDMS se produit à travers des trous traversants de 500 μm de diamètre. Après le cyclage thermique, le mélange réactionnel subit une purification dans la chambre de capture/purification, puis est injecté dans la chambre d'électrophorèse capillaire (CE). L'unité CE se compose d'un capillaire de 30 cm qui est plié en un motif de commutation compact via des spires de 65 μm de large.
Chimie de séquençage
- Cycle thermique
- Dans la chambre de réaction TC, le réactif de séquençage du terminateur de colorant, l'ADN modèle et les amorces sont chargés dans la chambre TC et soumis à un cycle thermique pendant 35 cycles (à 95 °C pendant 12 secondes et à 60 °C pendant 55 secondes).
- Purification
- Le mélange réactionnel chargé (contenant des fragments d'extension, un ADN modèle et un excès de réactif de séquençage) est conduit à travers une chambre de capture/purification à 30 °C via un champ électrique de 33 volts/cm appliqué entre les ports de sortie et d'entrée de capture. Le gel de capture à travers lequel l'échantillon est entraîné est constitué de 40 μM d'oligonucléotides (complémentaires aux amorces) liés de manière covalente à une matrice de polyacrylamide. Les fragments d'extension sont immobilisés par la matrice de gel et l'excès d'amorce, de modèle, de nucléotides libres et de sels sont élués à travers le port de déchets de capture. Le gel de capture est chauffé à 67-75 °C pour libérer les fragments d'extension.
- Électrophorèse capillaire
- Les fragments d'extension sont injectés dans la chambre CE où ils sont électrophorésés à travers un champ de 125-167 V/cm.
Plateformes
La plateforme Apollo 100 (Microchip Biotechnologies Inc., Dublin, CA) intègre les deux premières étapes du séquençage Sanger (cyclage thermique et purification) dans un système entièrement automatisé. Le fabricant affirme que les échantillons sont prêts pour l'électrophorèse capillaire dans les trois heures suivant le chargement de l'échantillon et des réactifs dans le système. La plateforme Apollo 100 nécessite des volumes de réactifs inférieurs au microlitre.
Comparaisons avec d'autres techniques de séquençage
L'objectif ultime du séquençage à haut débit est de développer des systèmes peu coûteux et extrêmement efficaces pour obtenir des longueurs de lecture étendues (plus longues). Des longueurs de lecture plus longues de chaque séparation électrophorétique unique réduisent considérablement le coût associé au séquençage de l'ADN de novo et le nombre de modèles nécessaires pour séquencer les contigs d'ADN à une redondance donnée. La microfluidique peut permettre un assemblage de séquences plus rapide, moins cher et plus facile.