
En écologie, une communauté est un groupe ou une association de populations de deux ou plusieurs espèces différentes occupant la même zone géographique au même moment, également appelée biocénose , communauté biotique , communauté biologique , communauté écologique ou assemblage de vie . Le terme communauté a de nombreuses utilisations. Dans sa forme la plus simple, il fait référence à des groupes d'organismes dans un lieu ou une période spécifique, par exemple, « la communauté de poissons du lac Ontario avant l'industrialisation ».
L'écologie des communautés ou synécologie est l'étude des interactions entre les espèces au sein des communautés à de nombreuses échelles spatiales et temporelles, y compris la distribution, la structure, l'abondance, la démographie et les interactions des populations coexistantes. L'écologie des communautés se concentre principalement sur les interactions entre les populations telles que déterminées par des caractéristiques génotypiques et phénotypiques spécifiques . Il est important de comprendre l'origine, le maintien et les conséquences de la diversité des espèces lors de l'évaluation de l'écologie des communautés.
L'écologie des communautés prend également en compte les facteurs abiotiques qui influencent la répartition ou les interactions des espèces (par exemple, la température annuelle ou le pH du sol ). Par exemple, les communautés végétales qui peuplent les déserts sont très différentes de celles que l'on trouve dans les forêts tropicales humides en raison des différences de précipitations annuelles. Les humains peuvent également affecter la structure des communautés en perturbant l'habitat , comme l'introduction d' espèces envahissantes .
À un niveau plus profond, la signification et la valeur du concept de communauté en écologie sont sujettes à débat. Les communautés ont traditionnellement été comprises à une échelle fine en termes de processus locaux construisant (ou détruisant) un assemblage d'espèces, comme la façon dont le changement climatique est susceptible d'affecter la composition des communautés d'herbes. Récemment, cette approche axée sur la communauté locale a été critiquée. Robert Ricklefs , professeur de biologie à l'Université du Missouri et auteur de Disintegration of the Ecological Community , a soutenu qu'il est plus utile de penser aux communautés à une échelle régionale, en s'appuyant sur la taxonomie évolutive et la biogéographie , où certaines espèces ou clades évoluent et d'autres disparaissent. Aujourd'hui, l'écologie des communautés se concentre sur les expériences et les modèles mathématiques, mais elle se concentrait auparavant principalement sur les modèles d'organismes. Par exemple, les subdivisions taxonomiques des communautés sont appelées populations , tandis que les partitions fonctionnelles sont appelées guildes .
Organisation
Niche
Au sein de la communauté, chaque espèce occupe une niche . La niche d'une espèce détermine la manière dont elle interagit avec l'environnement qui l'entoure et son rôle au sein de la communauté. En ayant différentes niches, les espèces peuvent coexister. C'est ce qu'on appelle la partition des niches. Par exemple, l'heure de la journée à laquelle une espèce chasse ou la proie qu'elle chasse.
Le partitionnement des niches réduit la concurrence entre les espèces de telle sorte que les espèces sont capables de coexister parce qu'elles suppriment leur propre croissance plus qu'elles ne limitent la croissance des autres espèces (c'est-à-dire que la concurrence au sein d'une espèce est supérieure à la concurrence entre les espèces, ou que la concurrence intraspécifique est supérieure à la concurrence interspécifique).
Le nombre de niches présentes dans une communauté détermine le nombre d'espèces présentes. Si deux espèces ont la même niche (par exemple, les mêmes besoins alimentaires), alors l'une des espèces prend le pas sur l'autre. Plus les niches sont remplies, plus la biodiversité de la communauté est élevée.
Niveau trophique
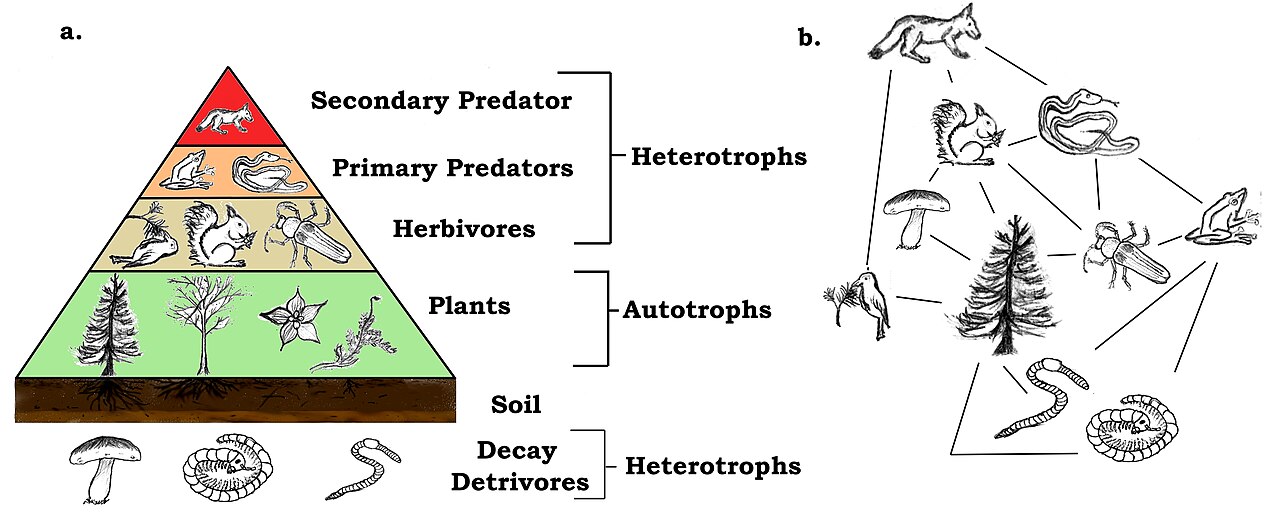
Le niveau trophique d'une espèce correspond à sa position dans la chaîne alimentaire ou le réseau. Au bas du réseau alimentaire se trouvent les autotrophes , également appelés producteurs primaires . Les producteurs fournissent leur propre énergie par photosynthèse ou chimiosynthèse , les plantes sont des producteurs primaires. Le niveau suivant est celui des herbivores (consommateurs primaires), ces espèces se nourrissent de végétation pour leur source d'énergie. Les herbivores sont consommés par les omnivores ou les carnivores . Ces espèces sont des consommateurs secondaires et tertiaires. Des niveaux supplémentaires à l'échelle trophique surviennent lorsque les omnivores ou les carnivores plus petits sont mangés par les plus gros. Au sommet du réseau alimentaire se trouve le prédateur au sommet , cette espèce animale n'est consommée par aucune autre espèce de la communauté. Les herbivores, les omnivores et les carnivores sont tous des hétérotrophes .
Un exemple de base de chaîne alimentaire est : herbe → lapin → renard. Les chaînes alimentaires deviennent plus complexes lorsque davantage d'espèces sont présentes, ce qui constitue souvent des réseaux alimentaires. L'énergie est transmise à travers les niveaux trophiques. L'énergie est perdue à chaque niveau, en raison d' inefficacités écologiques .
Le niveau trophique d'un organisme peut changer en fonction des autres espèces présentes. Par exemple, le thon peut être un prédateur au sommet de la chaîne alimentaire qui se nourrit de poissons plus petits, comme le maquereau. Cependant, dans une communauté où une espèce de requin est présente, le requin devient le prédateur au sommet de la chaîne alimentaire et se nourrit du thon.
Les décomposeurs jouent un rôle dans la pyramide trophique. Ils fournissent une source d'énergie et des nutriments aux espèces végétales de la communauté. Les décomposeurs tels que les champignons et les bactéries recyclent l'énergie vers la base du réseau trophique en se nourrissant d'organismes morts de tous les niveaux trophiques.
Guilde
Une guilde est un groupe d'espèces de la communauté qui utilisent les mêmes ressources de manière similaire. Les organismes d'une même guilde sont en compétition en raison de leur ressource commune. Des espèces étroitement apparentées font souvent partie de la même guilde, en raison de traits hérités par descendance commune de leur ancêtre commun . Cependant, les guildes ne sont pas exclusivement composées d'espèces étroitement apparentées.
Les carnivores, les omnivores et les herbivores sont tous des exemples de base de guildes. Une guilde plus précise serait celle des vertébrés qui se nourrissent d' arthropodes vivant au sol , ce qui comprendrait certains oiseaux et mammifères. Les plantes à fleurs qui ont le même pollinisateur forment également une guilde.
Espèces influentes
Certaines espèces ont une plus grande influence sur la communauté par leurs interactions directes et indirectes avec d’autres espèces. La population des espèces influentes est affectée par des perturbations abiotiques et biotiques. Ces espèces sont importantes pour identifier les communautés écologiques. La disparition de ces espèces entraîne de grands changements dans la communauté, réduisant souvent sa stabilité. Le changement climatique et l’introduction d’espèces envahissantes peuvent affecter le fonctionnement des espèces clés et donc avoir des répercussions sur les processus communautaires. L’industrialisation et l’introduction de polluants chimiques dans les environnements ont altéré à jamais les communautés et même des écosystèmes entiers.
Espèce fondatrice
Les espèces fondatrices influencent largement la population, la dynamique et les processus d'une communauté, en créant des changements physiques dans l'environnement lui-même. Ces espèces peuvent occuper n'importe quel niveau trophique, mais ont tendance à être productrices. palétuvier rouge est une espèce fondatrice des communautés marines. La racine de la mangrove fournit des zones de nurserie pour les jeunes poissons, tels que les vivaneaux .
Le pin à écorce blanche ( Pinus albicaulis ) est une espèce fondatrice. Après un incendie, l'arbre fournit de l'ombre (en raison de sa croissance dense), ce qui permet la repousse d'autres espèces végétales de la communauté. Cette croissance favorise le retour des invertébrés et des microbes nécessaires à la décomposition. Les graines de pin à écorce blanche fournissent de la nourriture aux grizzlis.

Espèce clé de voûte
Les espèces clés ont une influence disproportionnée sur la communauté par rapport à la plupart des espèces. Les espèces clés ont tendance à se situer aux niveaux trophiques supérieurs, étant souvent le prédateur au sommet. La suppression de l'espèce clé provoque des cascades trophiques descendantes . Les loups sont des espèces clés, étant un prédateur au sommet.
Dans le parc national de Yellowstone, la disparition de la population de loups due à la chasse excessive a entraîné une perte de biodiversité dans la communauté. Les loups contrôlaient le nombre d' élans dans le parc, par la prédation. Sans les loups, la population d'élans a considérablement augmenté, ce qui a entraîné un surpâturage. Cela a eu un impact négatif sur les autres organismes du parc ; le pâturage accru des élans a supprimé les sources de nourriture des autres animaux présents. Les loups ont depuis été réintroduits pour que la communauté du parc retrouve un fonctionnement optimal. Voir Réintroduction des loups et Histoire des loups à Yellowstone pour plus de détails sur cette étude de cas.
Un exemple marin d'espèce clé est Pisaster ochraceus . Cette étoile de mer contrôle l'abondance de Mytilus californianus , permettant ainsi de disposer de suffisamment de ressources pour les autres espèces de la communauté.
Ingénieurs écologiques
Un ingénieur écosystémique est une espèce qui maintient, modifie et crée des aspects d'une communauté. Il provoque des changements physiques dans l'habitat et altère les ressources disponibles pour les autres organismes présents.
Les castors qui construisent des barrages sont des ingénieurs écologiques. En coupant des arbres pour former des barrages, ils modifient le débit de l'eau dans une communauté. Ces changements influencent la végétation de la zone riveraine , et des études montrent que la biodiversité est accrue. Les terriers creusés par les castors créent des canaux, augmentant les connexions entre les habitats. Cela facilite le mouvement d'autres organismes de la communauté, comme les grenouilles.
Théories de la structure communautaire
La structure d'une communauté est sa composition. Elle est souvent mesurée à l'aide de réseaux biologiques , tels que les réseaux alimentaires . Les réseaux alimentaires sont une carte montrant les réseaux d'espèces et l'énergie qui relie les espèces entre elles par le biais d'interactions trophiques.
Théorie holistique
La théorie holistique fait référence à l'idée selon laquelle une communauté est définie par les interactions entre les organismes qui la composent. Toutes les espèces sont interdépendantes, chacune jouant un rôle essentiel dans le fonctionnement de la communauté. De ce fait, les communautés sont répétables et faciles à identifier, avec des facteurs abiotiques similaires qui les contrôlent.
Frédéric Clements a développé le concept holistique (ou organismique) de communauté, comme s'il s'agissait d'un superorganisme ou d'une unité discrète, avec des limites nettes. Clements a proposé cette théorie après avoir remarqué que certaines espèces de plantes se trouvaient régulièrement ensemble dans des habitats, il a conclu que les espèces étaient dépendantes les unes des autres. La formation des communautés n'est pas aléatoire et implique une coévolution .
La théorie holistique découle de la pensée plus large du holisme , qui fait référence à un système composé de nombreuses parties, toutes nécessaires au fonctionnement du système.
Théorie individualiste
Henry Gleason a développé le concept individualiste (également connu sous le nom de concept ouvert ou continuum) de communauté, dans lequel l'abondance d'une population d'une espèce change progressivement le long de gradients environnementaux complexes. Chaque espèce change indépendamment par rapport aux autres espèces présentes le long du gradient. L'association des espèces est aléatoire et due à la coïncidence. Les conditions environnementales variables et la probabilité de chaque espèce d'arriver et de s'établir le long du gradient influencent la composition de la communauté.
La théorie individualiste propose que les communautés puissent exister en tant qu’entités continues, en plus des groupes discrets mentionnés dans la théorie holistique.
Théorie neutre
Stephen P. Hubbell a introduit la théorie neutre de l'écologie (à ne pas confondre avec la théorie neutre de l'évolution moléculaire ). Au sein de la communauté (ou métacommunauté ), les espèces sont fonctionnellement équivalentes et l'abondance d'une population d'une espèce change par des processus démographiques stochastiques (c'est-à-dire des naissances et des décès aléatoires). L'équivalence des espèces dans la communauté conduit à une dérive écologique. La dérive écologique conduit à des populations d'espèces fluctuant de manière aléatoire, tandis que le nombre total d'individus dans la communauté reste constant. Lorsqu'un individu meurt, il y a une chance égale que chaque espèce colonise cette parcelle. Les changements stochastiques peuvent entraîner l'extinction d'espèces au sein de la communauté, mais cela peut prendre beaucoup de temps s'il y a de nombreux individus de cette espèce.
Les espèces peuvent coexister parce qu'elles sont similaires, les ressources et les conditions appliquent un filtre au type d'espèces présentes dans la communauté. Chaque population a la même valeur adaptative (capacités de compétition et de dispersion) et la même demande en ressources. La composition locale et régionale représente un équilibre entre la spéciation ou la dispersion (qui augmentent la diversité) et les extinctions aléatoires (qui diminuent la diversité).
Interactions interspécifiques
Les espèces interagissent de diverses manières : compétition, prédation , parasitisme , mutualisme , commensalisme , etc. L'organisation d'une communauté biologique en fonction des interactions écologiques est appelée structure communautaire.
Concours
Les espèces peuvent entrer en compétition les unes avec les autres pour des ressources limitées . On considère que cette compétition est un facteur limitant important de la taille de la population , de la biomasse et de la richesse spécifique . De nombreux types de compétition ont été décrits, mais la preuve de l'existence de ces interactions est sujette à débat. Une compétition directe a été observée entre individus, populations et espèces, mais il existe peu de preuves que la compétition ait été la force motrice de l'évolution des grands groupes.
- Compétition d'interférence : se produit lorsqu'un individu d'une espèce interfère directement avec un individu d'une autre espèce. Cela peut se faire pour de la nourriture ou pour un territoire. On peut citer comme exemple un lion qui chasse une hyène après l'avoir tuée, ou une plante qui libère des substances chimiques allélopathiques pour entraver la croissance d'une espèce concurrente.
- Compétition apparente : se produit lorsque deux espèces partagent un prédateur. Par exemple, un couguar chasse le caribou des bois et le cerf. Les populations des deux espèces peuvent être réduites par la prédation sans concurrence d'exploitation directe.
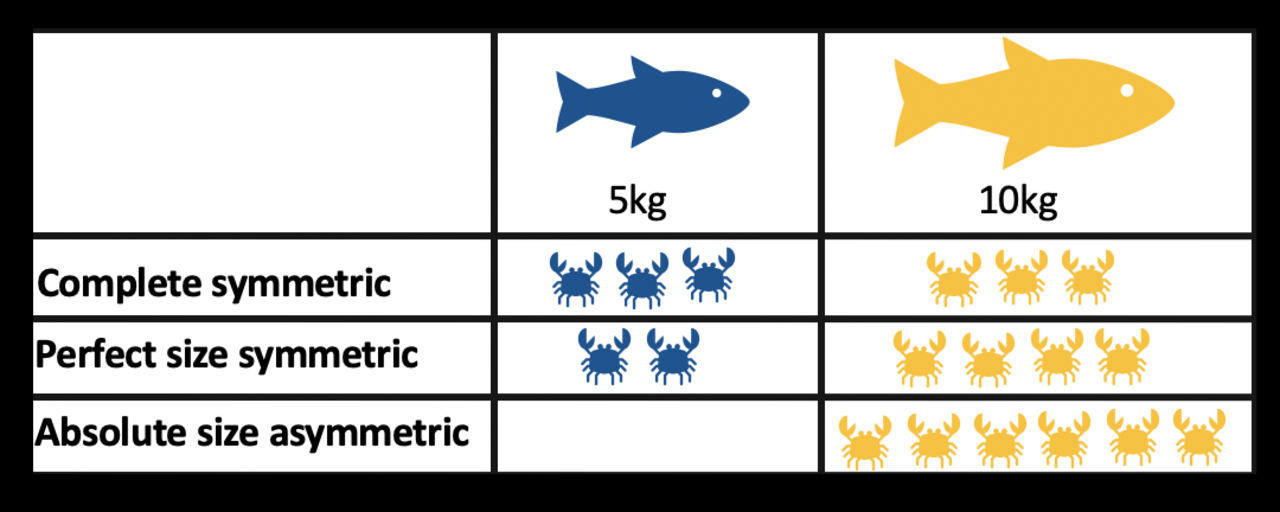
- Compétition d'exploitation : elle se produit par la consommation de ressources. Lorsqu'un individu d'une espèce consomme une ressource (par exemple, de la nourriture, un abri, la lumière du soleil, etc.), cette ressource n'est plus disponible pour la consommation par un membre d'une autre espèce. On pense que la compétition d'exploitation est plus courante dans la nature, mais il faut veiller à la distinguer de la compétition apparente. Un exemple de compétition d'exploitation pourrait être celui entre des herbivores consommant de la végétation ; le lapin et le cerf mangent tous deux de l'herbe des prés. La compétition d'exploitation varie :
- complètement symétrique - tous les individus reçoivent la même quantité de ressources, quelle que soit leur taille
- taille parfaite symétrique - tous les individus exploitent la même quantité de ressources par unité de biomasse
- taille absolue asymétrique - les plus grands individus exploitent toutes les ressources disponibles.
- Le degré d’asymétrie de taille a des effets majeurs sur la structure et la diversité des communautés écologiques
Prédation
La prédation consiste à chasser une autre espèce pour se nourrir. Il s'agit d'une interaction positive-négative, l'espèce prédatrice en profite tandis que l'espèce proie est lésée. Certains prédateurs tuent leur proie avant de la manger, ce qu'on appelle aussi tuer et consommer. Par exemple, un faucon attrape et tue une souris. D'autres prédateurs sont des parasites qui se nourrissent de proies vivantes, par exemple une chauve-souris vampire se nourrissant d'une vache. Le parasitisme peut cependant entraîner la mort de l'organisme hôte au fil du temps. Un autre exemple est l'alimentation des plantes d' herbivores , par exemple une vache qui broute. L'herbivorie est un type de prédation dans lequel une plante (la proie dans cet exemple) tentera de dissuader le prédateur de manger la plante en injectant une toxine dans les feuilles de la plante. Cela peut amener le prédateur à consommer d'autres zones de la plante ou à ne pas consommer la plante du tout. La prédation peut affecter la taille de la population de prédateurs et de proies et le nombre d'espèces coexistant dans une communauté.
La prédation peut être spécialisée, par exemple la belette pygmée chasse uniquement le campagnol des champs. Ou généraliste, par exemple l'ours polaire mange principalement des phoques mais peut changer son régime alimentaire pour se tourner vers les oiseaux lorsque la population de phoques est faible.
Les espèces peuvent être des prédateurs solitaires ou en groupe. L'avantage de chasser en groupe est qu'il est possible de capturer des proies plus grosses, mais la source de nourriture doit être partagée. Les loups sont des prédateurs en groupe, tandis que les tigres sont des prédateurs solitaires.
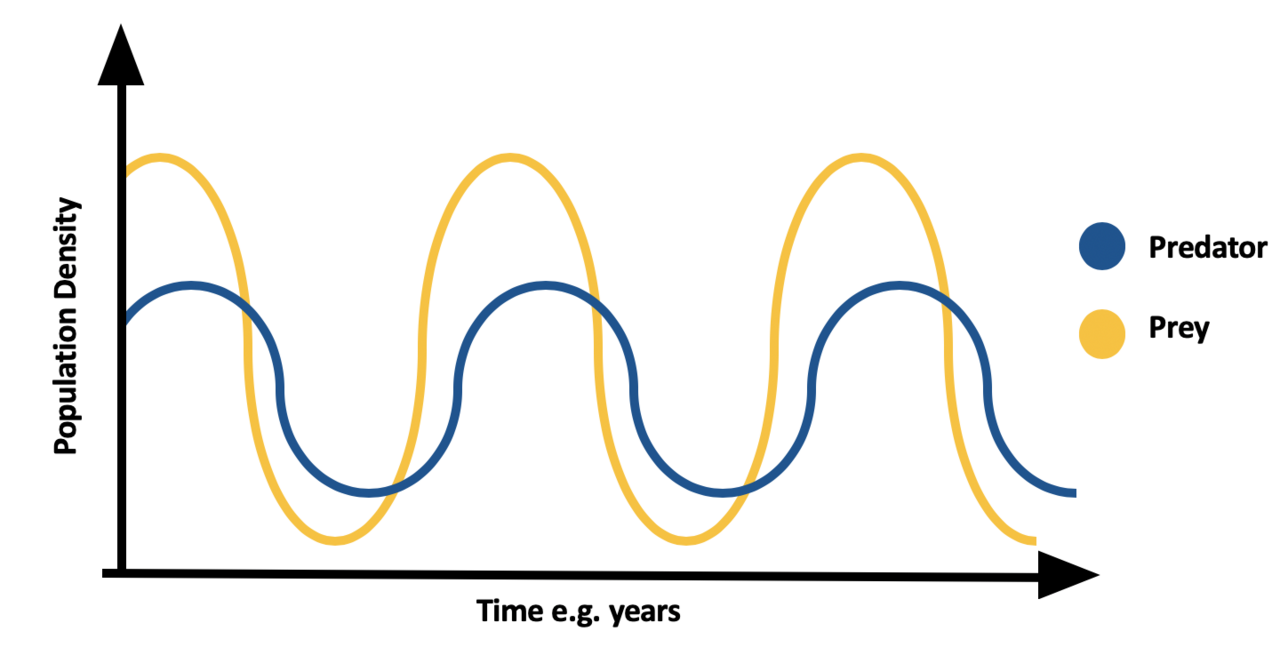
La prédation dépend de la densité , ce qui conduit souvent à des cycles de population. Lorsque les proies sont abondantes, les espèces prédatrices augmentent, mangeant ainsi davantage d'espèces proies et provoquant le déclin de la population de proies. En raison du manque de nourriture, la population de prédateurs diminue. En raison du manque de prédation, la population de proies augmente. Voir les équations de Lotka-Volterra pour plus de détails à ce sujet. Un exemple bien connu de cela est celui des cycles de population lynx - lièvre observés dans le nord.
La prédation peut entraîner une coévolution : une course aux armements évolutive , la proie s'adapte pour éviter le prédateur, le prédateur évolue. Par exemple, une espèce proie développe une toxine qui tue son prédateur et le prédateur développe une résistance à la toxine, la rendant ainsi inoffensive.
Mutualisme
Le mutualisme est une interaction entre espèces dans laquelle les deux espèces bénéficient.
Un exemple est celui des bactéries Rhizobium qui se développent dans des nodules sur les racines des légumineuses. Cette relation entre la plante et les bactéries est endosymbiotique , les bactéries vivant sur les racines de la légumineuse. La plante fournit aux bactéries des composés fabriqués lors de la photosynthèse, qui peuvent être utilisés comme source d'énergie. Alors que Rhizobium est une bactérie fixatrice d'azote , fournissant des acides aminés ou de l'ammonium à la plante.
Les insectes pollinisant les fleurs des angiospermes en sont un autre exemple. De nombreuses plantes dépendent de la pollinisation par un pollinisateur. Un pollinisateur transfère le pollen de la fleur mâle au stigmate de la fleur femelle . Cela fertilise la fleur et permet à la plante de se reproduire. Les abeilles, comme les abeilles domestiques , sont les pollinisateurs les plus connus. Les abeilles obtiennent du nectar de la plante qu'elles utilisent comme source d'énergie. Le pollen non transféré fournit des protéines à l'abeille. La plante bénéficie de la fertilisation, tandis que l'abeille reçoit de la nourriture.
Commensalisme
Le commensalisme est un type de relation entre organismes dans lequel un organisme bénéficie tandis que l'autre organisme n'est ni avantagé ni lésé. L'organisme qui bénéficie est appelé le commensal tandis que l'autre organisme qui n'est ni avantagé ni lésé est appelé l' hôte .
Par exemple, une orchidée épiphyte attachée à l'arbre pour la soutenir est bénéfique pour l'orchidée, mais ne lui nuit ni ne lui profite. Ce type de commensalisme est appelé inquilinisme , l'orchidée vit en permanence sur l'arbre.
La phorésie est un autre type de commensalisme, le commensal utilise l'hôte uniquement pour le transport. De nombreuses espèces d'acariens dépendent d'un autre organisme, comme les oiseaux ou les mammifères, pour se disperser.
La métabiose est la forme finale du commensalisme. Le commensal dépend de l'hôte pour préparer un environnement propice à la vie. Par exemple, le varech possède un système de type racine, appelé crampon , qui le fixe au fond marin. Une fois enraciné, il fournit aux mollusques , comme les escargots de mer, un abri qui les protège de la prédation.
Amensalisme
L'opposé du commensalisme est l'amensalisme , une relation interspécifique dans laquelle un produit d'un organisme a un effet négatif sur un autre organisme, mais l'organisme d'origine n'est pas affecté.
Un exemple est l'interaction entre les têtards de la grenouille commune et un escargot d'eau douce . Les têtards consomment de grandes quantités de microalgues. Les algues étant moins abondantes pour l'escargot, les algues disponibles pour l'escargot sont également de moindre qualité. Le têtard a donc un effet négatif sur l'escargot sans en tirer un avantage notable. Les têtards obtiendraient la même quantité de nourriture avec ou sans la présence de l'escargot.
Un arbre plus vieux et plus grand peut inhiber la croissance des arbres plus petits. Un jeune arbre poussant à l'ombre d'un arbre mature a du mal à obtenir de la lumière pour la photosynthèse. L'arbre mature a également un système racinaire bien développé, ce qui l'aide à concurrencer le jeune arbre pour les nutriments. La croissance du jeune arbre est donc entravée, ce qui entraîne souvent sa mort. La relation entre les deux arbres est l'amensalisme, l'arbre mature n'étant pas affecté par la présence du plus petit.
Parasitisme
Le parasitisme est une interaction dans laquelle un organisme, l’hôte, est lésé tandis que l’autre, le parasite, en bénéficie.
Le parasitisme est une symbiose , un lien à long terme dans lequel le parasite se nourrit de l'hôte ou prend des ressources de celui-ci. Les parasites peuvent vivre à l'intérieur du corps, comme le ténia . Ou à la surface du corps, comme les poux de tête .

Le paludisme résulte d'une relation parasitaire entre un moustique femelle du genre Anopheles et Plasmodium . Les moustiques attrapent le parasite en se nourrissant d'un vertébré infecté. À l'intérieur du moustique, le plasmodium se développe dans la paroi de l'intestin moyen. Une fois développé en zygote, le parasite se déplace vers les glandes salivaires où il peut être transmis à une espèce de vertébré, par exemple l'homme. Le moustique agit comme vecteur du paludisme. Le parasite a tendance à réduire la durée de vie du moustique et inhibe la production de progéniture.
Un deuxième exemple de parasitisme est le parasitisme de couvée . Les coucous pratiquent régulièrement ce type de parasitisme. Les coucous pondent leurs œufs dans le nid d'une autre espèce d'oiseau. L'hôte s'occupe donc du poussin du coucou comme s'il s'agissait du sien, sans pouvoir faire la différence. Les poussins du coucou éjectent les petits de l'hôte du nid, ce qui signifie qu'ils bénéficient d'un niveau de soins et de ressources plus important de la part des parents. L'élevage des jeunes est coûteux et peut réduire le succès de la progéniture future, c'est pourquoi le coucou tente d'éviter ce coût en parasitant la couvée.
De la même manière que la prédation, le parasitisme peut conduire à une course aux armements évolutive . L'hôte évolue pour se protéger du parasite et le parasite évolue pour surmonter cette restriction.
Neutralisme
Le neutralisme est une situation dans laquelle les espèces interagissent, mais cette interaction n'a aucun effet notable sur l'une ou l'autre des espèces concernées. En raison de l'interdépendance des communautés, le véritable neutralisme est rare. Les exemples de neutralisme dans les systèmes écologiques sont difficiles à prouver, en raison des effets indirects que les espèces peuvent avoir les unes sur les autres.