Les plumes sont des excroissances épidermiques qui forment un revêtement externe distinctif, ou plumage , chez les oiseaux et certains dinosaures non aviens ainsi que chez d'autres archosaures . Elles constituent les structures tégumentaires les plus complexes chez les vertébrés et un exemple de nouveauté évolutive complexe . Elles figurent parmi les caractéristiques qui distinguent les oiseaux actuels des autres groupes vivants
Bien que les plumes recouvrent la majeure partie du corps de l'oiseau, elles ne poussent que sur certaines zones bien définies de la peau. Elles contribuent au vol, à l'isolation thermique et à l'imperméabilité. De plus, la coloration facilite la communication et la protection . L'étude des plumes s'appelle la plumologie (ou science du plumage ).
Les plumes sont utilisées de multiples façons, tant pratiques que culturelles et religieuses. Douces et excellentes pour retenir la chaleur , elles sont parfois employées dans la literie haut de gamme , notamment pour les oreillers , les couvertures et les matelas . Elles servent également de garnissage pour les vêtements d'hiver et la literie d'extérieur, comme les manteaux matelassés et les sacs de couchage . Le duvet d'oie et d'eider possède un gonflant exceptionnel , c'est-à-dire la capacité de se déployer à partir d'un état comprimé et stocké pour emprisonner de grandes quantités d'air isolant. Les plumes de grands oiseaux (le plus souvent d'oies ) ont été et sont encore utilisées pour fabriquer des plumes d'oie . Historiquement, la chasse aux oiseaux pour leurs plumes décoratives a mis en danger certaines espèces et a contribué à l'extinction d'autres. Aujourd'hui, les plumes utilisées dans la mode et pour les coiffures et vêtements militaires proviennent des déchets de l'élevage de volailles, notamment de poulets , d'oies , de dindes , de faisans et d'autruches . Ces plumes sont teintes et travaillées pour en améliorer l'apparence, car les plumes de volaille sont souvent naturellement plus ternes que celles des oiseaux sauvages.
Les plumes d'oie , premiers instruments d'écriture, étaient fabriquées à partir de plumes. Le mot « plume » lui-même dérive du latin « penna » , qui signifie plume. Le mot français « plume » peut désigner une plume , une plume d'oie ou un stylo .
Structures et caractéristiques
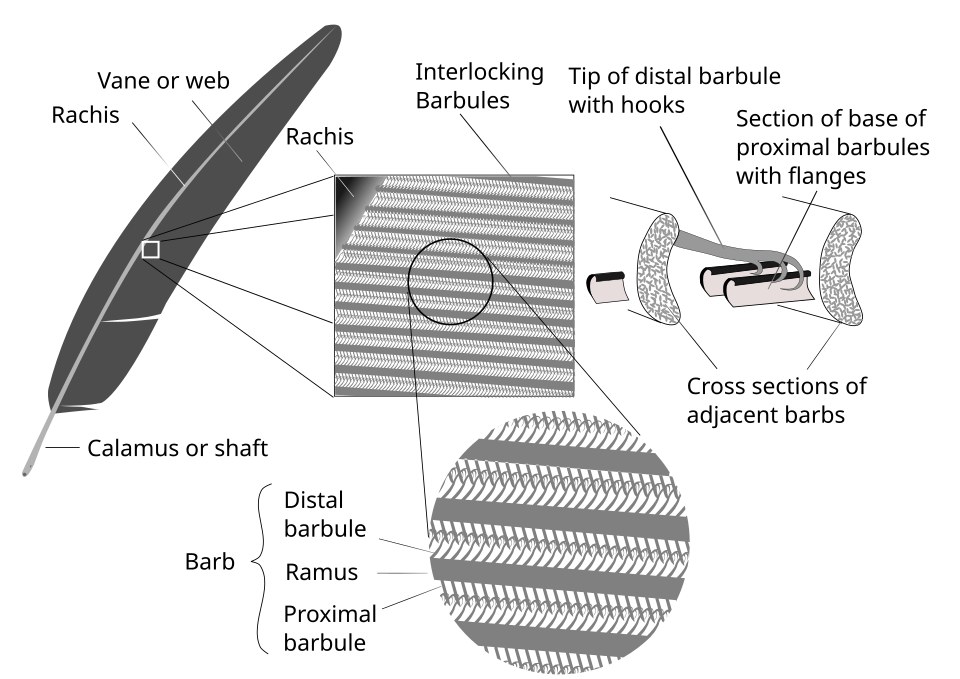

Les plumes figurent parmi les appendices tégumentaires les plus complexes des vertébrés. Elles se forment dans de minuscules follicules de l' épiderme , ou couche externe de la peau, qui produisent des protéines de kératine . Les β-kératines des plumes, des becs et des griffes – ainsi que des griffes, des écailles et des carapaces des reptiles – sont composées de chaînes protéiques liées par des liaisons hydrogène en feuillets β plissés . Ces feuillets sont ensuite torsadés et réticulés par des ponts disulfure pour former des structures encore plus résistantes que les α-kératines des poils , des cornes et des sabots des mammifères . Les signaux précis qui induisent la croissance des plumes sur la peau restent inconnus, mais il a été démontré que le facteur de transcription cDermo-1 induit la croissance des plumes sur la peau et des écailles sur les pattes.
Classification

Il existe deux types principaux de plumes : les plumes à barbe, qui recouvrent l’extérieur du corps, et le duvet , situé sous les plumes à barbe. Les plumes pennées sont des plumes à barbe. Également appelées plumes de contour, elles prennent naissance à partir de bandes et recouvrent l’ensemble du corps. Un troisième type de plume, plus rare, le filoplume , est semblable à un poil et étroitement associé aux plumes pennées. Il est souvent entièrement dissimulé par celles-ci ; un ou deux filoplumes sont attachés et poussent à proximité du même point de la peau que chaque plume pennée, du moins sur la tête, le cou et le tronc de l’oiseau. Les filoplumes sont totalement absents chez les ratites . Chez certains passereaux , des filoplumes apparaissent au-delà des plumes pennées sur le cou. Les rémiges, ou rémiges de l’aile, et les rectrices, ou rémiges de la queue, sont les plumes les plus importantes pour le vol. Une plume typique à barbes présente un rachis principal . Des ramifications, ou barbes , sont soudées au rachis ; ces barbes sont elles-mêmes ramifiées et forment les barbules . Ces barbules sont munies de minuscules crochets, les barbicelles, permettant leur fixation croisée. Les plumes de duvet sont duveteuses car elles sont dépourvues de barbicelles ; les barbules flottent donc librement, ce qui permet au duvet d'emprisonner l'air et d'offrir une excellente isolation thermique. À la base de la plume, le rachis s'élargit pour former le calamus (ou rachis ), un tube creux qui s'insère dans un follicule cutané . La partie basale du calamus est dépourvue de barbes. Cette partie est enchâssée dans le follicule cutané et présente une ouverture à sa base (ombilic proximal) et une petite ouverture latérale (ombilic distal).
Les oisillons de certaines espèces possèdent un type particulier de duvet natal (néossoptiles) qui sont expulsés lorsque les plumes normales (téléoptiles) émergent.
Les rémiges sont rigides afin de résister à l'air lors du battement descendant, mais restent souples dans les autres directions. On a observé que l'orientation des fibres de β-kératine dans les plumes des oiseaux volants diffère de celle des oiseaux incapables de voler : les fibres sont mieux alignées le long de l'axe de la tige vers l'extrémité , et les parois latérales du rachis présentent une structure de fibres entrecroisées
Fonctions
Les plumes isolent les oiseaux de l'eau et du froid. Elles peuvent également être arrachées pour tapisser le nid et isoler les œufs et les oisillons. Les plumes des ailes et de la queue jouent un rôle important dans le contrôle du vol. Certaines espèces possèdent une crête de plumes sur la tête. Bien que les plumes soient légères, le plumage d'un oiseau pèse deux à trois fois plus que son squelette, car de nombreux os sont creux et contiennent des sacs aériens. Les motifs colorés servent de camouflage aux oiseaux dans leur habitat et aux prédateurs en quête de nourriture. Comme chez les poissons, la coloration du dessus et du dessous du corps peut différer pour assurer le camouflage en vol. Les différences marquées dans les motifs et les couleurs des plumes font partie du dimorphisme sexuel de nombreuses espèces d'oiseaux et sont particulièrement importantes dans le choix des partenaires. Dans certains cas, on observe des différences de réflectivité UV des plumes entre les sexes, même si aucune différence de couleur n'est visible dans le spectre visible. Les plumes des ailes des manakins à ailes massues mâles Machaeropterus deliciosus possèdent des structures spéciales qui sont utilisées pour produire des sons par stridulation .

Certains oiseaux possèdent un duvet poudreux qui se renouvelle continuellement, de petites particules se détachant régulièrement de l'extrémité des barbules. Ces particules produisent une poudre qui se répartit entre les plumes du corps de l'oiseau et agit comme agent imperméabilisant et conditionneur . Ce duvet poudreux a évolué indépendamment chez plusieurs taxons et se trouve aussi bien dans le duvet que dans les plumes pennées. Il peut être dispersé dans le plumage, comme chez les pigeons et les perroquets, ou concentré en plaques localisées sur la poitrine, le ventre ou les flancs, comme chez les hérons et les podarges. Les hérons utilisent leur bec pour briser le duvet poudreux et le répartir, tandis que les cacatoès peuvent utiliser leur tête comme une houppette pour l'appliquer. L'imperméabilité peut être altérée par l'exposition à des agents émulsifiants dus à la pollution humaine. Les plumes peuvent alors se gorger d'eau, ce qui peut entraîner la noyade de l'oiseau. Il est également très difficile de nettoyer et de secourir les oiseaux dont le plumage a été souillé par des marées noires . Les plumes des cormorans absorbent l'eau et contribuent à réduire la flottabilité, permettant ainsi aux oiseaux de nager sous l'eau.

Les soies sont des plumes rigides et effilées, dotées d'un rachis large mais de peu de barbes. Les soies rictales se trouvent autour des yeux et du bec. Elles pourraient avoir une fonction similaire à celle des cils et des vibrisses chez les mammifères . Bien qu'il n'existe pas encore de preuves formelles, il a été suggéré que les soies rictales possèdent des fonctions sensorielles et pourraient aider les oiseaux insectivores à capturer leurs proies. Dans une étude, il a été constaté que les gobemouches des saules ( Empidonax traillii ) capturaient les insectes aussi bien avant qu'après l'ablation des soies rictales.
Les grèbes sont particuliers par leur habitude d'ingérer leurs propres plumes et de les donner à leurs petits. Les observations sur leur régime alimentaire composé de poissons et la fréquence de l'ingestion de plumes suggèrent que l'ingestion de plumes, en particulier du duvet de leurs flancs, contribue à la formation de pelotes facilement éjectables.
Distribution
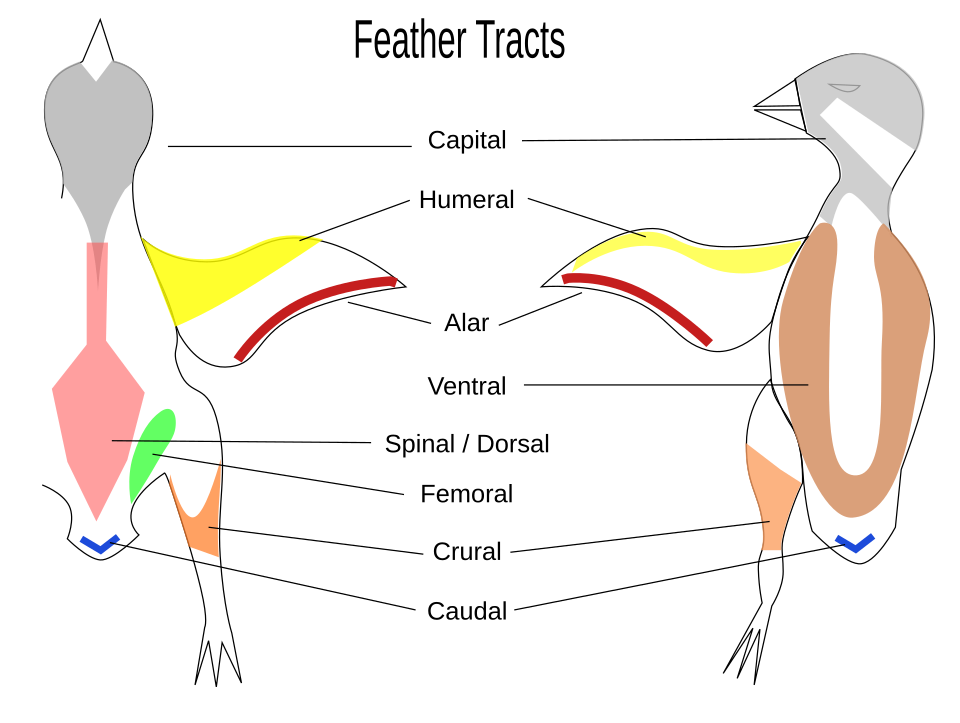
Les plumes de contour ne sont pas réparties uniformément sur la peau des oiseaux, sauf chez certains groupes comme les manchots , les ratites et les kamichis. Chez la plupart des oiseaux , les plumes poussent à partir de zones cutanées spécifiques appelées ptéryles ; entre les ptéryles se trouvent des régions dépourvues de plumes, appelées aptères . Les filoplumes et le duvet peuvent provenir des aptères. La disposition de ces zones de plumes, la ptérylose ou ptérylographie, varie selon les familles d’oiseaux et a été utilisée par le passé pour déterminer les relations évolutives entre ces familles. Les espèces qui couvent leurs propres œufs perdent souvent leurs plumes sur une partie de leur ventre, formant une plaque de couvage .
Coloration

La couleur des plumes est produite par des pigments, par des structures microscopiques capables de réfracter , de réfléchir ou de diffuser certaines longueurs d'onde de la lumière, ou par une combinaison des deux.
La plupart des pigments des plumes sont des mélanines ( phéomélanines brunes et beiges , eumélanines noires et grises ) et des caroténoïdes (rouges, jaunes, oranges) ; d'autres pigments n'apparaissent que chez certains taxons – les psittacofulvines jaunes à rouges (présentes chez certains perroquets ) et la turacine rouge et la turacoverdine verte ( pigments porphyriniques que l'on trouve uniquement chez les touracos ).
La coloration structurale intervient dans la production des couleurs bleues, l'irisation , la majeure partie de la réflectance ultraviolette et dans l'intensification des couleurs pigmentaires. Une irisation structurale a été observée dans des plumes fossiles datant de 40 millions d'années. Les plumes blanches, dépourvues de pigment, diffusent la lumière ; l'albinisme chez les oiseaux est dû à une production défectueuse de pigments, sans que la coloration structurale ne soit affectée (comme on peut l'observer, par exemple, chez les perruches ondulées bleues et blanches ).

Les teintes bleues et vertes éclatantes de nombreux perroquets résultent d'interférences constructives de la lumière réfléchie par différentes couches de structures au sein des plumes. Dans le cas du plumage vert, outre le jaune, la structure spécifique de la plume impliquée est parfois appelée texture de Dyck. La mélanine participe souvent à l'absorption de la lumière ; combinée à un pigment jaune, elle produit un vert olive terne.

Chez certains oiseaux, la couleur du plumage peut être créée ou modifiée par les sécrétions de la glande uropygienne , également appelée glande uropygienne. La couleur jaune du bec de nombreux calaos est due à ces sécrétions. Il a été suggéré qu'il existe d'autres différences de couleur, visibles uniquement dans l'ultraviolet , mais les études n'ont pas permis de les confirmer La sécrétion huileuse de la glande uropygienne pourrait également avoir un effet inhibiteur sur les bactéries présentes sur les plumes
Les couleurs rouges, oranges et jaunes de nombreuses plumes sont dues à divers caroténoïdes. Les pigments à base de caroténoïdes pourraient être des signaux fiables de santé, car ils proviennent de régimes alimentaires spécifiques et sont donc parfois difficiles à obtenir , et/ou parce que les caroténoïdes sont nécessaires au fonctionnement du système immunitaire et que, par conséquent, les parades nuptiales se font au détriment de la santé
Les plumes d'un oiseau s'usent et sont remplacées périodiquement au cours de sa vie par la mue . Les nouvelles plumes, appelées plumes de sang ou plumes en formation selon leur stade de développement, se forment à partir des mêmes follicules que les anciennes. La présence de mélanine dans les plumes augmente leur résistance à l'abrasion. Une étude note que les plumes riches en mélanine se dégradent plus rapidement sous l'action bactérienne, même comparées aux plumes non pigmentées de la même espèce, que celles non pigmentées ou contenant des pigments caroténoïdes. Cependant, une autre étude, menée la même année, a comparé l'action des bactéries sur la pigmentation de deux espèces de bruants chanteurs et a observé que les plumes les plus foncées étaient plus résistantes ; les auteurs ont cité d'autres recherches, également publiées en 2004, qui indiquaient qu'une plus grande quantité de mélanine conférait une meilleure résistance. Ils ont observé que la plus grande résistance des oiseaux aux plumes plus foncées confirmait la règle de Gloger .
Bien que la sélection sexuelle joue un rôle majeur dans le développement du plumage, et notamment dans sa couleur, ce n'est pas la seule explication. De nouvelles études suggèrent que le plumage unique des oiseaux influence fortement de nombreux aspects importants de leur comportement, comme la hauteur à laquelle différentes espèces construisent leurs nids. Les femelles étant les principales responsables des soins aux petits, l'évolution a favorisé la sélection de femelles arborant un plumage aux couleurs plus ternes, leur permettant de se fondre dans l'environnement du nid. La position du nid et le risque accru de prédation ont également influencé le plumage des femelles. Une espèce nichant au sol, plutôt que dans la canopée, devra présenter des couleurs beaucoup plus ternes pour ne pas attirer l'attention sur son nid. L'étude sur la hauteur de nidification a révélé que les oiseaux nichant dans la canopée subissent souvent davantage d'attaques de prédateurs en raison de la couleur plus vive du plumage des femelles. Une autre influence de l'évolution qui pourrait expliquer la grande variété de couleurs et de motifs des plumes d'oiseaux est que ces dernières ont développé leurs couleurs vives grâce à la végétation et aux fleurs environnantes. Les oiseaux développent leurs couleurs vives en vivant au milieu de certaines couleurs. La plupart des espèces d'oiseaux se fondent dans leur environnement grâce à un certain degré de camouflage. Ainsi, si l'habitat d'une espèce est riche en couleurs et en motifs, l'espèce évoluera pour se camoufler et éviter d'être la proie des oiseaux. Les plumes d'oiseaux présentent une vaste gamme de couleurs, surpassant même la variété de couleurs de nombreuses plantes, feuilles et fleurs.
Plumes utilisées lors des parades nuptiales
Chez les oiseaux présentant un dimorphisme sexuel, les mâles développent souvent une coloration distincte ou des plumes ornementales spécialisées utilisées lors des parades nuptiales pour attirer les femelles. Plusieurs théories ont été proposées quant à l'origine de ces plumes ornementales, les premiers exemples ayant été observés chez plusieurs théropodes primitifs (voir la sous-section « Origines » ci-dessous).
L'exemple le plus connu de plumage ornemental utilisé lors de la parade nuptiale est celui du paon mâle ( Pavo cristatus) . Les mâles arborent une longue traîne de plumes couvertes, ornée de motifs ocelles distinctifs, qu'ils mettent en valeur lors d'une parade vigoureuse. Au cours de ces parades, les mâles déploient leur traîne en éventail, exhibant ainsi leur plumage aux femelles.
L'origine évolutive du plumage ornemental et des parades nuptiales du paon demeure incertaine, plusieurs théories proposant une combinaison de facteurs. Dans une étude observant le comportement de parade des paons, la longueur de leur traîne, la longueur de leur torse et la densité de leurs ocelles ont été mesurées chez des paons mâles en captivité. Ces derniers ont ensuite été placés dans des enclos avec des paonnes, et leur succès reproductif a été mesuré par le nombre de copulations réussies. Les résultats ont montré que le choix des femelles n'était pas influencé par la longueur de la traîne, mais par la densité des ocelles. Ceci suggère que le plumage élaboré et les parades nuptiales des paons mâles ont évolué en fonction du choix des femelles, qui privilégieraient particulièrement les mâles présentant une plus grande diversité d'ocelles.
Structure et utilisation
La morphologie osseuse du radius, du cubitus et de l'humérus, qui soutiennent les plumes ornementales, peut également influencer le choix des femelles. Par exemple, la morphologie osseuse des mâles du manakin à ailes massues ( Machaeropterus deliciosus ) est très spécialisée : leurs cubitus sont plus grands et plus denses, avec un volume et une densité minérale supérieurs à ceux des autres membres de la famille des manakins. Les rémiges secondaires sont hypertrophiées et servent lors des parades nuptiales ; les mâles entrechoquent l'extrémité de ces rémiges de façon répétée, produisant un son caractéristique. La parade se termine par un déploiement rapide des longues plumes jaunes de la gorge, accompagné de sauts.
Longueur de la queue

La longueur de la queue est un autre exemple de l'influence des plumes sur le choix du partenaire, comme chez le veuve à longue queue ( Euplectes progne ). Les mâles de cette espèce arborent des plumes caudales extrêmement longues pendant la saison des amours, ce qui est corrélé à un plus grand succès reproductif. Dans une étude célèbre testant la corrélation entre la longueur de la queue et la valeur sélective , la longueur de la queue de mâles de veuves à longue queue a été artificiellement raccourcie ou allongée ; ceux à queue allongée ont présenté un meilleur succès reproductif. Il a également été proposé que les plumes ornementales plus longues jouent un rôle dans la défense du territoire et la compétition intrasexuelle, comme moyen d'affirmer sa dominance, mais aucune preuve substantielle n'est venue étayer cette théorie.
Origine des plumes ornementales
L'une des origines possibles des plumes ornementales se trouve chez le mégalosauroïde Sciurumimus , qui présente une morphologie simple et monofilamenteuse. Les plumes monofilamenteuses possèdent une structure unique, semblable à un fil, contrairement aux plumes ramifiées ou barbelées, et sont considérées comme la forme la plus ancienne de plumes. Des plumes monofilamenteuses ont également été découvertes chez un large éventail de taxons, bien que Sciurumimus soit le plus ancien. On en a également trouvé chez le tyrannosauroïde Yutyrannus et le thérizinosauroïde Beipiaosaurus , avec des monofilaments proportionnellement plus larges qui constituaient probablement une forme primitive de plumes ornementales spécialisées.
L’analyse de plumes découvertes dans de l’ambre birman a révélé une coloration inhabituelle le long du rachis , suggérant qu’elles arboraient des motifs de couleurs remarquables. Les premières plumes ornementales du genre Schizooura suggèrent une fonction à la fois aérodynamique et ornementale, leur forme en queue d’épingle étant trop étroite pour avoir un impact sur l’aérodynamisme.
Parasites
La surface des plumes abrite certains ectoparasites, notamment les poux des plumes ( Pthiraptera ) et les acariens des plumes. Les poux des plumes vivent généralement sur un seul hôte et ne peuvent se transmettre que des parents aux poussins, entre oiseaux en période de reproduction et, occasionnellement, par phorésie . Ce cycle de vie explique que la plupart des espèces de parasites soient spécifiques à leur hôte et coévoluent avec lui, ce qui les rend intéressantes pour les études phylogénétiques.
Les trous dans les plumes sont des traces de rongement causées par des poux (très probablement des poux du genre Brueelia ) sur les plumes des ailes et de la queue. Ils ont été décrits chez l'hirondelle rustique et, du fait de leur facilité de comptage, de nombreuses publications en évolution, écologie et comportementologie les utilisent pour quantifier l'intensité de l'infestation.
Les coucous parasites qui grandissent dans les nids d'autres espèces ont également des poux de plumes spécifiques à l'hôte et ceux-ci semblent être transmis seulement après que les jeunes coucous aient quitté le nid de l'hôte.
Les oiseaux entretiennent leur plumage en le lissant et en se baignant dans l'eau ou la poussière . Il a été suggéré qu'un comportement particulier des oiseaux, le fourmiliage , au cours duquel des fourmis sont introduites dans le plumage, contribue à réduire les parasites, mais aucune preuve à l'appui n'a été trouvée.
Utilisation humaine
Utilitaire


Les plumes d'oiseaux sont utilisées depuis longtemps pour empenner les flèches . Les plumes colorées, comme celles des faisans, servent à décorer les leurres de pêche .
Les plumes sont également précieuses pour l'identification des espèces dans le cadre d'études médico-légales, notamment en cas de collisions d'oiseaux avec des aéronefs. Les rapports isotopiques de l'hydrogène dans les plumes permettent de déterminer l'origine géographique des oiseaux. Les plumes peuvent aussi être utiles pour l'échantillonnage non destructif des polluants.
L'industrie avicole produit une grande quantité de plumes, qui, comme d'autres formes de kératine, se décomposent lentement. Ces déchets de plumes ont été utilisés dans plusieurs applications industrielles : comme milieu de culture pour les micro-organismes , pour la production de polymères biodégradables et d'enzymes . Les protéines de plumes ont également été testées comme adhésif pour panneaux de bois
Certains groupes autochtones d'Alaska utilisent des plumes de lagopède comme dégraissant (additifs non plastiques) dans la fabrication de poteries depuis le premier millénaire avant J.-C. afin de promouvoir la résistance aux chocs thermiques et la solidité.
En matière de religion et de culture

Les plumes d'aigle revêtent une grande valeur culturelle et spirituelle pour les Amérindiens des États-Unis et les Premières Nations du Canada , où elles sont considérées comme des objets religieux. Aux États-Unis, l'usage religieux des plumes d'aigle et de faucon est régi par la loi fédérale sur les plumes d'aigle , qui limite leur possession aux membres certifiés et inscrits des tribus amérindiennes reconnues par le gouvernement fédéral.
En Amérique du Sud, des décoctions à base de plumes de condor sont utilisées en médecine traditionnelle. En Inde, les plumes du paon indien sont utilisées en médecine traditionnelle pour traiter les morsures de serpent, l'infertilité et la toux.
Les membres du clan Campbell d'Écosse sont connus pour porter des plumes sur leur coiffe, symbole d'autorité au sein du clan. Les chefs de clan en portent trois, les chefs de clan deux et un armiger une. Tout membre du clan ne répondant pas à ces critères n'est pas autorisé à porter de plumes dans le cadre de sa tenue traditionnelle, et ce faisant, il est considéré comme présomptueux.
Aux XVIIIe, XIXe et début du XXe siècles, un commerce international florissant de plumes pour la confection de chapeaux et autres coiffes féminines extravagantes (notamment à la mode victorienne ) connut un essor considérable. En 1886, Frank Chapman constata que les plumes de près de 40 espèces d'oiseaux étaient utilisées dans environ les trois quarts des 700 chapeaux de dames qu'il observa à New York. Par exemple, on utilisait autrefois des plumes de colibri d'Amérique du Sud pour habiller certains des oiseaux miniatures présents dans les nichoirs à oiseaux chanteurs . Ce commerce entraîna de graves pertes pour les populations d'oiseaux (notamment les aigrettes et les grues blanches d'Amérique ).
Les défenseurs de l'environnement ont mené des campagnes contre l'utilisation des plumes dans les chapeaux, contribuant ainsi à d'importantes mesures de protection de l'environnement et à l'évolution de la mode. On peut citer en exemple la création de la Royal Society for the Protection of Birds au Royaume-Uni en 1889, ou l'adoption du Lacey Act aux États-Unis en 1900. Le marché des plumes ornementales s'est alors largement effondré.
Plus récemment, le plumage du coq est devenu une tendance populaire comme accessoire de coiffure , les plumes autrefois utilisées comme leurres de pêche étant désormais utilisées pour apporter couleur et style aux cheveux.
La production d'articles en plumes en Europe a décliné ces 60 dernières années, principalement en raison de la concurrence asiatique. Les plumes ont longtemps orné les chapeaux lors d'événements prestigieux tels que les mariages et la Journée des Dames aux courses hippiques (Royal Ascot).
Évolution


Considérations fonctionnelles
La vision fonctionnelle de l'évolution des plumes s'est traditionnellement concentrée sur l'isolation, le vol et la parade nuptiale. Cependant, la découverte de dinosaures à plumes non volants du Crétacé supérieur en Chine suggère que le vol ne pouvait être leur fonction première, car les plumes n'auraient tout simplement pas été capables de fournir la moindre portance Il a été suggéré que les plumes auraient pu avoir pour fonction première la thermorégulation, l'imperméabilisation, voire l'élimination des déchets métaboliques tels que le soufre . Des découvertes récentes semblent confirmer l'hypothèse d'une fonction thermorégulatrice, du moins chez les petits dinosaures . Certains chercheurs avancent même que la thermorégulation serait apparue grâce aux soies faciales utilisées comme capteurs tactiles . Bien que l'hypothèse d'une évolution des plumes à partir d' écailles de reptiles ait été avancée , cette idée fait l'objet de nombreuses objections, et des explications plus récentes s'inscrivent dans le paradigme de la biologie évolutive du développement . Les théories sur l'origine des plumes à partir des écailles suggèrent que la structure plane de ces dernières s'est modifiée pour se développer en plumes par division afin de former la membrane. Cependant, ce processus de développement implique une structure tubulaire issue d'un follicule, ce tube se divisant longitudinalement pour former la membrane. Le nombre de plumes par unité de surface cutanée est plus élevé chez les petits oiseaux que chez les grands, ce qui souligne leur rôle important dans l'isolation thermique. En effet, les petits oiseaux perdent davantage de chaleur en raison de leur surface corporelle relativement plus importante par rapport à leur poids. La miniaturisation des oiseaux a également joué un rôle dans l'évolution du vol actif. On pense que la coloration des plumes a évolué principalement en réponse à la sélection sexuelle . Chez les spécimens fossiles du paravien Anchiornis huxleyi et du ptérosaure Tupandactylus imperator , les caractéristiques sont si bien conservées que la structure des mélanosomes (cellules pigmentaires) est observable. En comparant la forme des mélanosomes fossiles à celle des mélanosomes d'oiseaux actuels, il a été possible de déterminer la couleur et le motif des plumes d' Anchiornis et de Tupandactylus Anchiornisprésentait un plumage à motifs noirs et blancs sur les membres antérieurs et postérieurs, avec une crête brun rougeâtre. Ce motif est similaire à la coloration de nombreuses espèces d'oiseaux actuelles, qui utilisent la coloration du plumage pour la parade nuptiale et la communication, notamment pour la sélection sexuelle et le camouflage. Il est probable que les espèces de dinosaures non aviens utilisaient des motifs de plumage pour des fonctions similaires à celles des oiseaux modernes avant l'apparition du vol. Dans de nombreux cas, l'état physiologique des oiseaux (en particulier des mâles) est indiqué par la qualité de leurs plumes, et cette caractéristique est utilisée par les femelles pour lechoix du partenaire. De plus, en comparant différents spécimens d'Ornithomimus edmontonicus , on a constaté que les individus plus âgés possédaient un pennibrachium (une structure en forme d'aile composée de plumes allongées), contrairement aux plus jeunes. Cela suggère que le pennibrachium était un caractère sexuel secondaire et qu'il avait probablement une fonction sexuelle.
Évolution moléculaire
Plusieurs gènes déterminant le développement des plumes ont été identifiés. Ils seront essentiels à la compréhension de l'évolution des plumes. Par exemple, certains gènes transforment les écailles en plumes ou en structures semblables à des plumes lorsqu'ils sont exprimés ou induits dans les pattes des oiseaux, comme les convertisseurs écaille-plume Sox2 , Zic1 , Grem1 , Spry2 et Sox18 .
Les plumes et les écailles sont composées de deux formes distinctes de kératine , et l'on a longtemps cru que chaque type de kératine était spécifique à chaque structure cutanée (plumes et écailles). Cependant, la kératine des plumes est également présente aux premiers stades de développement des écailles de l'alligator d'Amérique . Ce type de kératine, que l'on pensait auparavant spécifique aux plumes, est inhibé au cours du développement embryonnaire de l'alligator et est donc absent des écailles des alligators adultes. La présence de cette kératine homologue chez les oiseaux et les crocodiliens indique qu'elle a été héritée d'un ancêtre commun.
Ceci pourrait suggérer que les écailles de crocodiliens, les plumes d'oiseaux et d'autres dinosaures, ainsi que les pycnofibres de ptérosaures , sont toutes des expressions développementales des mêmes structures cutanées primitives d'archosaures ; suggérant ainsi que les plumes et les pycnofibres pourraient être homologues. Des méthodes de datation moléculaire utilisées en 2011 montrent que la sous-famille des β-kératines de plumes présentes chez les oiseaux actuels a commencé à diverger il y a 143 millions d'années, suggérant que les plumes pennées d' Anchiornis n'étaient pas composées des β-kératines de plumes présentes chez les oiseaux actuels. Cependant, une étude de plumes fossiles du dinosaure Sinosauropteryx et d'autres fossiles a révélé des traces de protéines en feuillets β, grâce à la spectroscopie infrarouge et à la spectroscopie des rayons X du soufre. La présence abondante de protéines α dans certaines plumes fossiles s'est avérée être un artefact du processus de fossilisation, car les structures des protéines β se transforment facilement en hélices α lors de la dégradation thermique. En 2019, des scientifiques ont découvert que les gènes responsables de la production de plumes ont évolué à la base des archosaures, confirmant ainsi que les plumes étaient présentes chez les premiers ornithodires et concordant avec les données fossiles.
Dinosaures à plumes
Plusieurs dinosaures non aviens possédaient des plumes sur leurs membres qui ne leur permettaient pas de voler. Une théorie suggère que les plumes sont apparues chez les dinosaures grâce à leurs propriétés isolantes ; puis, les petites espèces de dinosaures qui ont développé des plumes plus longues les auraient trouvées utiles pour planer, ce qui aurait conduit à l’évolution de proto-oiseaux comme l’Archaeopteryx et le Microraptor zhaoianus . Une autre théorie postule que l’avantage adaptatif initial des premières plumes résidait dans leur pigmentation ou leur irisation, contribuant à la préférence sexuelle dans le choix du partenaire. Parmi les dinosaures possédant des plumes ou des proto-plumes, on trouve le Pedopenna daohugouensis et le Dilong paradoxus , un tyrannosauroïde qui est de 60 à 70 millions d’années plus ancien que le Tyrannosaurus rex .
La majorité des dinosaures connus pour avoir possédé des plumes ou des proto-plumes sont des théropodes . Cependant, des structures tégumentaires filamenteuses semblables à des plumes ont également été observées chez les ornithischiens Tianyulong et Psittacosaurus [ La nature exacte de ces structures est encore à l'étude. On pense néanmoins que les plumes de stade 1 (voir la section « Stades évolutifs » ci-dessous), telles que celles observées chez ces deux ornithischiens, avaient probablement une fonction de parade . En 2014, l'ornithischien Kulindadromeus a été décrit comme possédant des structures ressemblant à des plumes de stade 3 . L'hypothèse d'une évolution des écailles chez les premiers ancêtres des dinosaures est élevée. Toutefois, cette hypothèse repose sur le postulat que les ptérosaures primitifs étaient recouverts d'écailles. Une étude de 2016 analyse la morphologie pulpaire des soies caudales du Psittacosaurus et constate qu'elles sont similaires à des plumes, mais aussi aux soies de la tête du paon du Congo , à la barbe du dindon sauvage et à l'épine crânienne du kamikaze . Une réestimation par maximum de vraisemblance réalisée par le paléontologue Thomas Holtz suggère que les filaments étaient plus probablement l'état ancestral des dinosaures.
En 2010, la présence de rémiges sur le cubitus d'un carcharodontosauridé nommé Concavenator corcovatus a été découverte , suggérant la présence de structures ressemblant à des plumes sur ses bras . Cependant, Foth et al. (2014) contestent cette hypothèse, soulignant que les protubérances présentes sur le cubitus de Concavenator sont situées sur sa face antérolatérale , contrairement aux rémiges postérolatérales observées chez certains oiseaux. Ils estiment plus probable qu'il s'agisse de points d'insertion pour des ligaments interosseux . Cette hypothèse a été réfutée par Cuesta Fidalgo et ses collègues, qui ont également constaté que ces protubérances sont postérolatérales, contrairement à la position des ligaments interosseux
Depuis les années 1990, des dizaines de dinosaures à plumes ont été découverts dans le clade des Maniraptora , qui comprend le clade des Avialae et les ancêtres communs récents des oiseaux, les Oviraptorosauria et les Deinonychosauria . En 1998, la découverte d'un oviraptorosaurien à plumes, Caudipteryx zoui , a remis en question l'idée que les plumes étaient une structure exclusive aux Avialae. Enfoui dans la formation d'Yixian, au Liaoning (Chine), C. zoui a vécu au Crétacé inférieur. Présentes sur les membres antérieurs et la queue, ses plumes ont été identifiées Troodontidae et les Dromaeosauridae . Des plumes ramifiées avec rachis, barbes et barbules ont été découvertes chez de nombreux membres, y compris Sinornithosaurus millenii , un droméosauridé trouvé dans la formation d'Yixian (124,6 millions d'années).
Auparavant, un paradoxe temporel existait dans l'évolution des plumes : les théropodes présentant des caractéristiques aviaires très développées sont apparus plus tard qu'Archaeopteryx , suggérant que les descendants des oiseaux étaient apparus avant l'ancêtre commun. Cependant, la découverte d' Anchiornis huxleyi dans la formation de Tiaojishan (Jurassique supérieur, 160 millions d'années) dans l'ouest du Liaoning en 2009 a résolu ce paradoxe. Antérieur à Archaeopteryx , Anchiornis prouve l'existence d'un ancêtre théropode à plumes modernes, offrant un éclairage nouveau sur la transition dinosaures-oiseaux. Le spécimen présente une répartition de grandes plumes pennées sur les membres antérieurs et la queue, ce qui implique que les plumes pennées se sont étendues au reste du corps à un stade plus précoce de l'évolution des théropodes . Le développement des plumes pennées n'a pas remplacé les plumes filamenteuses antérieures. Des plumes filamenteuses sont conservées aux côtés de plumes de vol d'apparence moderne – dont certaines avec des modifications que l'on retrouve dans les plumes des oiseaux plongeurs actuels – dans de l'ambre vieux de 80 millions d'années provenant de l'Alberta.
Deux petites ailes piégées dans de l'ambre datant de 100 millions d'années montrent que le plumage existait chez certains ancêtres des oiseaux. Ces ailes appartenaient très probablement aux énantiornithes , un groupe diversifié de dinosaures aviens.
Une vaste analyse phylogénétique des premiers dinosaures, réalisée par Matthew Baron, David B. Norman et Paul Barrett (2017), a révélé que les Théropodes sont en réalité plus étroitement apparentés aux Ornithischiens , dont ils forment le groupe frère au sein du clade des Ornithoscelida . Cette étude suggère également que si les structures plumeuses des théropodes et des ornithischiens ont une origine évolutive commune, alors il est possible que les plumes soient apparues exclusivement chez les Ornithoscelida. Dans ce cas, l'apparition des plumes remonterait probablement au Trias moyen [ bien que cette hypothèse soit contestée L'absence de plumes chez les grands sauropodes et les ankylosaures pourrait s'expliquer par la suppression de leur développement par des régulateurs génomiques
stades évolutifs

Plusieurs études sur le développement des plumes chez les embryons d'oiseaux modernes, associées à la répartition des types de plumes chez divers ancêtres préhistoriques des oiseaux, ont permis aux scientifiques de tenter de reconstituer la séquence dans laquelle les plumes ont d'abord évolué pour donner naissance aux types que l'on trouve chez les oiseaux modernes.
L'évolution des plumes a été décomposée en les étapes suivantes par Xu et Guo en 2009 :
- filament unique
- Plusieurs filaments reliés à leur base
- Plusieurs filaments reliés à leur base à un filament central
- Plusieurs filaments répartis le long d'un filament central
- Plusieurs filaments émergeant du bord d'une structure membranaire
- Plume pennée à barbes et barbules et rachis central
- Plume pennée à rachis asymétrique
- Aile indifférenciée à rachis central
Cependant, Foth (2011) a montré que certains de ces stades supposés (les stades 2 et 5 en particulier) sont probablement de simples artefacts de conservation dus à la manière dont les plumes fossilisées sont écrasées et dont les restes ou empreintes de plumes sont préservés. Foth a réinterprété les plumes du stade 2 comme étant des plumes écrasées ou mal identifiées d'au moins le stade 3, et les plumes du stade 5 comme étant des plumes écrasées du stade 6.
Le diagramme simplifié suivant, illustrant les relations phylogénétiques des dinosaures, fait suite à ces résultats et présente la répartition probable des plumes duveteuses et pennées chez les dinosaures et les oiseaux préhistoriques. Ce diagramme est basé sur celui présenté par Xu et Guo (2009) , modifié à partir des travaux de Foth (2011) . Les chiffres accompagnant chaque nom indiquent la présence de différents stades de développement du plumage. La lettre « s » signale la présence d'écailles sur le corps.
Chez les ptérosaures
On savait depuis longtemps que les ptérosaures possédaient des structures filamenteuses ressemblant à de la fourrure, appelées pycnofibres , recouvrant leur corps. Ces structures étaient généralement considérées comme distinctes des « vraies plumes » des oiseaux et des dinosaures apparentés. Cependant, une étude menée en 2018 sur deux petits fossiles de ptérosaures bien conservés, provenant du Jurassique de Mongolie-Intérieure (Chine) , a révélé que les ptérosaures étaient recouverts d'un ensemble de pycnofibres de structures variées (et non seulement filamenteuses). Plusieurs de ces structures présentaient des caractéristiques diagnostiques des plumes, telles que des filaments groupés sans nervures et des filaments ramifiés bilatéralement, deux caractéristiques que l'on pensait initialement exclusives aux oiseaux et aux autres dinosaures maniraptoriens. Au vu de ces découvertes, il est possible que les plumes aient des origines évolutives profondes chez les archosaures ancestraux , bien qu'il soit également possible que ces structures aient évolué indépendamment pour ressembler aux plumes d'oiseaux par le biais d'une évolution convergente . Mike Benton, principal auteur de l'étude, a appuyé la première théorie, déclarant : « Nous n'avons trouvé aucune preuve anatomique que les quatre types de pycnofibres diffèrent des plumes des oiseaux et des dinosaures. Par conséquent, puisqu'ils sont identiques, ils doivent partager une origine évolutive commune, qui remonte à environ 250 millions d'années, bien avant l'apparition des oiseaux. » Cependant, comme l'a souligné Liliana D'Alba, la structure tégumentaire des spécimens d' anurognathidés repose toujours sur la morphologie générale. Les pycnofibres des deux spécimens d'anurognathidés pourraient ne pas être homologues aux appendices filamenteux des dinosaures. Paul M. Barrett suppose que, lors de l'évolution tégumentaire des ptérosaures, ces derniers ont perdu primitivement leurs écailles et que des pycnofibres ont commencé à apparaître.
Cascocauda était presque entièrement recouvert d'un épais pelage de pycnofibres, qui semblent avoir été de deux types. Le premier type est constitué de filaments simples et incurvés, mesurant de 3,5 à 12,8 mm de long. Ces filaments recouvrent la majeure partie de l'animal, y compris la tête, le cou, le corps, les membres et la queue. Le second type est composé de touffes de filaments soudés près de leur base, semblables aux plumes duveteuses ramifiées des oiseaux et autres dinosaures cœlurosauriens , mesurant environ 2,5 à 8,0 mm de long et ne recouvrant que les membranes alaires. L'étude d'échantillons de pycnofibres a révélé la présence de microcorps au sein des filaments, ressemblant aux pigments mélanosomiques identifiés dans d'autres téguments fossiles, notamment les phéomélanosomes. De plus, l'analyse spectrale infrarouge de ces pycnofibres montre des spectres d'absorption similaires à ceux des cheveux roux humains . Ces pycnofibres ont probablement assuré l'isolation thermique et contribué à l'aérodynamisme du corps et des ailes en vol. L'identité de ces structures ramifiées, à savoir des pycnofibres ou des plumes, a été remise en question par Unwin et Martill (2020), qui les ont interprétées comme des aktinofibrilles dégradées et regroupées – des fibres rigidifiantes présentes dans la membrane alaire des ptérosaures – et ont attribué les mélanosomes et la kératine à la peau plutôt qu'à des filaments. Ces affirmations ont été réfutées par Yang et ses collègues, qui soutiennent que les interprétations d'Unwin et Martill sont incompatibles avec l'état de conservation du spécimen. Plus précisément, ils affirment que la structure constante, l'espacement régulier et l'extension des filaments au-delà de la membrane alaire confirment leur identification comme pycnofibres. De plus, ils soutiennent que la présence exclusive de mélanosomes et de kératine dans les fibres, comme c'est le cas pour les plumes de dinosaures fossiles, confirme qu'il s'agit de filaments et exclut une contamination par de la peau conservée. Les proto-plumes ont probablement évolué chez les premiers archosaures, peu de temps après l'extinction du PT, au moment où les taux métaboliques des premiers archosaures et synapsides augmentaient, les postures devenaient érigées et l'activité soutenue.