Un organisme multicellulaire est un organisme constitué de plusieurs cellules , contrairement aux organismes unicellulaires . Toutes les espèces d' animaux , de plantes terrestres et la plupart des champignons sont multicellulaires, tout comme de nombreuses algues , tandis que quelques organismes sont partiellement unicellulaires et partiellement multicellulaires, comme les moisissures visqueuses et les amibes sociales telles que le genre Dictyostelium .
Les organismes multicellulaires apparaissent de diverses manières, par exemple par division cellulaire ou par agrégation de nombreuses cellules individuelles. Les organismes coloniaux sont le résultat de la réunion de nombreux individus identiques pour former une colonie . Cependant, il peut souvent être difficile de distinguer les protistes coloniaux des véritables organismes multicellulaires, car les deux concepts ne sont pas distincts ; les protistes coloniaux ont été surnommés « pluricellulaires » plutôt que « multicellulaires ». Il existe également des organismes macroscopiques qui sont multinucléés bien que techniquement unicellulaires, comme les Xénophyophores qui peuvent atteindre 20 cm.
Histoire évolutive
Occurrence
Français La multicellularité a évolué indépendamment au moins 25 fois chez les eucaryotes , et aussi chez certains procaryotes , comme les cyanobactéries , les myxobactéries , les actinomycètes , Magnetoglobus multicellularis ou Methanosarcina . Cependant, les organismes multicellulaires complexes n'ont évolué que dans six groupes eucaryotes : les animaux , les champignons symbiomycètes , les algues brunes , les algues rouges , les algues vertes et les plantes terrestres . Elle a évolué à plusieurs reprises pour les Chloroplastida (algues vertes et plantes terrestres), une fois pour les animaux, une fois pour les algues brunes, trois fois pour les champignons ( chytrides , ascomycètes et basidiomycètes ) et peut-être plusieurs fois pour les moisissures visqueuses et les algues rouges. La première preuve d'une organisation multicellulaire, qui se produit lorsque des organismes unicellulaires coordonnent leurs comportements et peut être un précurseur évolutif de la véritable multicellularité, provient d' organismes de type cyanobactérie qui ont vécu il y a 3,0 à 3,5 milliards d'années. Pour se reproduire, les véritables organismes multicellulaires doivent résoudre le problème de la régénération d'un organisme entier à partir de cellules germinales (c'est-à-dire des spermatozoïdes et des ovules ), un problème qui est étudié en biologie du développement évolutif . Les animaux ont développé une diversité considérable de types de cellules dans un corps multicellulaire (100 à 150 types de cellules différents), contre 10 à 20 chez les plantes et les champignons.
Perte de multicellularité
La perte de multicellularité s'est produite dans certains groupes. Les champignons sont principalement multicellulaires, bien que les premières lignées divergentes soient en grande partie unicellulaires (par exemple, Microsporidia ) et qu'il y ait eu de nombreux retours à l'unicellularité chez les champignons (par exemple, Saccharomycotina , Cryptococcus et d'autres levures ). Elle peut également s'être produite chez certaines algues rouges (par exemple, Porphyridium ), mais elles peuvent être primitivement unicellulaires. La perte de multicellularité est également considérée comme probable chez certaines algues vertes (par exemple, Chlorella vulgaris et certaines Ulvophyceae ). Dans d'autres groupes, généralement des parasites, une réduction de la multicellularité s'est produite, dans le nombre ou les types de cellules (par exemple, les myxozoaires , des organismes multicellulaires, que l'on pensait auparavant unicellulaires, sont probablement des cnidaires extrêmement réduits ).
Cancer
Les organismes multicellulaires, en particulier les animaux à longue durée de vie, sont confrontés au défi du cancer , qui survient lorsque les cellules ne parviennent pas à réguler leur croissance dans le cadre du programme normal de développement. Des changements dans la morphologie des tissus peuvent être observés au cours de ce processus. Le cancer chez les animaux ( métazoaires ) a souvent été décrit comme une perte de multicellularité et une réversion atavique vers un état de type unicellulaire. De nombreux gènes responsables de l'établissement de la multicellularité qui ont pris naissance autour de l'apparition des métazoaires sont dérégulés dans les cellules cancéreuses, y compris les gènes qui contrôlent la différenciation cellulaire , l'adhésion et la communication intercellulaire . Il existe une discussion sur la possibilité de l'existence du cancer dans d'autres organismes multicellulaires ou même dans les protozoaires . Par exemple, les galles des plantes ont été caractérisées comme des tumeurs , mais certains auteurs soutiennent que les plantes ne développent pas de cancer.
Séparation des cellules somatiques et germinales
Dans certains groupes multicellulaires, appelés weismannistes , une séparation entre une lignée cellulaire somatique stérile et une lignée cellulaire germinale a évolué. Cependant, le développement weismanniste est relativement rare (par exemple, les vertébrés, les arthropodes, Volvox ), car une grande partie des espèces ont la capacité d' embryogenèse somatique (par exemple, les plantes terrestres, la plupart des algues, de nombreux invertébrés).
Hypothèses d'origine
Une hypothèse sur l'origine de la multicellularité est qu'un groupe de cellules à fonction spécifique se sont agrégées en une masse en forme de limace appelée grex , qui se déplaçait comme une unité multicellulaire. C'est essentiellement ce que font les moisissures visqueuses . Une autre hypothèse est qu'une cellule primitive a subi une division du noyau, devenant ainsi un cœnocyte . Une membrane se formerait alors autour de chaque noyau (et de l'espace cellulaire et des organites occupés dans l'espace), ce qui donnerait naissance à un groupe de cellules connectées dans un seul organisme (ce mécanisme est observable chez la drosophile ). Une troisième hypothèse est que lorsqu'un organisme unicellulaire se divisait, les cellules filles ne parvenaient pas à se séparer, ce qui donnait lieu à une conglomération de cellules identiques dans un seul organisme, qui pourrait plus tard développer des tissus spécialisés. C'est ce que font les embryons végétaux et animaux ainsi que les choanoflagellés coloniaux .
Les premiers organismes multicellulaires étant des organismes simples et mous, dépourvus d'os, de coquille ou d'autres parties dures du corps, ils ne sont pas bien conservés dans les archives fossiles. Une exception pourrait être la démosponge , qui a peut-être laissé une signature chimique dans les roches anciennes. Les premiers fossiles d'organismes multicellulaires comprennent le Grypania spiralis contesté et les fossiles des schistes noirs de la formation fossile B du groupe Francevillien du Paléoprotérozoïque au Gabon ( Gabonionta ). La formation de Doushantuo a livré des microfossiles vieux de 600 millions d'années avec des preuves de traits multicellulaires.
Jusqu'à récemment, la reconstruction phylogénétique se faisait par le biais de similitudes anatomiques (en particulier embryologiques ). Cette hypothèse est inexacte, car les organismes multicellulaires vivants tels que les animaux et les plantes sont éloignés de leurs ancêtres unicellulaires de plus de 500 millions d'années. Un tel écoulement du temps permet à la fois aux évolutions divergentes et convergentes d'imiter les similitudes et d'accumuler les différences entre les groupes d'espèces ancestrales modernes et éteintes. La phylogénétique moderne utilise des techniques sophistiquées telles que les alloenzymes , l'ADN satellite et d'autres marqueurs moléculaires pour décrire les traits partagés par des lignées éloignées.
L’évolution de la multicellularité aurait pu se produire de plusieurs manières différentes, dont certaines sont décrites ci-dessous :
La théorie symbiotique
Cette théorie suggère que les premiers organismes multicellulaires sont nés de la symbiose (coopération) de différentes espèces d'organismes unicellulaires, chacune ayant des rôles différents. Au fil du temps, ces organismes seraient devenus si dépendants les uns des autres qu'ils ne seraient plus capables de survivre indépendamment, ce qui a finalement conduit à l'incorporation de leurs génomes dans un seul organisme multicellulaire. Chaque organisme respectif deviendrait une lignée distincte de cellules différenciées au sein de l'espèce nouvellement créée.
Ce type de symbiose fortement co-dépendante peut être observé fréquemment, comme dans la relation entre les poissons clowns et les anémones de mer Riterri . Dans ces cas, il est extrêmement douteux que l'une ou l'autre espèce puisse survivre très longtemps si l'autre venait à disparaître. Cependant, le problème avec cette théorie est qu'on ne sait toujours pas comment l'ADN de chaque organisme pourrait être incorporé dans un seul génome pour les constituer en une seule espèce. Bien que l'on suppose qu'une telle symbiose s'est produite (par exemple, les mitochondries et les chloroplastes dans les cellules animales et végétales - endosymbiose ), elle ne s'est produite que très rarement et, même dans ce cas, les génomes des endosymbiotes ont conservé un élément de distinction, en répliquant séparément leur ADN pendant la mitose de l'espèce hôte. Par exemple, les deux ou trois organismes symbiotiques formant le lichen composite , bien que dépendants les uns des autres pour leur survie, doivent se reproduire séparément puis se reformer pour créer à nouveau un organisme individuel.
La théorie de la cellularisation (syncytiale)
Cette théorie stipule qu'un seul organisme unicellulaire, avec plusieurs noyaux , aurait pu développer des cloisons membranaires internes autour de chacun de ses noyaux. De nombreux protistes tels que les ciliés ou les moisissures visqueuses peuvent avoir plusieurs noyaux, ce qui étaye cette hypothèse . Cependant, la simple présence de plusieurs noyaux ne suffit pas à étayer la théorie. Les noyaux multiples des ciliés sont différents et ont des fonctions clairement différenciées. Le macro -noyau répond aux besoins de l'organisme, tandis que le micro- noyau est utilisé pour la reproduction sexuée avec échange de matériel génétique. Les syncitia des moisissures visqueuses se forment à partir de cellules amiboïdes individuelles, comme les tissus syncitiaux de certains organismes multicellulaires, et non l'inverse. Pour être jugée valide, cette théorie a besoin d'un exemple démontrable et d'un mécanisme de génération d'un organisme multicellulaire à partir d'un syncytium préexistant.
La théorie coloniale
La théorie coloniale de Haeckel , 1874, propose que la symbiose de nombreux organismes de la même espèce (contrairement à la théorie symbiotique, qui suggère la symbiose d'espèces différentes) a conduit à un organisme multicellulaire. Au moins une certaine multicellularité - on suppose qu'elle est issue de l'évolution terrestre - se produit lorsque des cellules se séparent puis se rejoignent (par exemple, les moisissures visqueuses cellulaires ), alors que pour la majorité des types multicellulaires (ceux qui ont évolué dans des environnements aquatiques), la multicellularité se produit en conséquence de l'incapacité des cellules à se séparer après la division. Le mécanisme de cette dernière formation de colonie peut être aussi simple qu'une cytokinèse incomplète , bien que la multicellularité soit également généralement considérée comme impliquant une différenciation cellulaire .
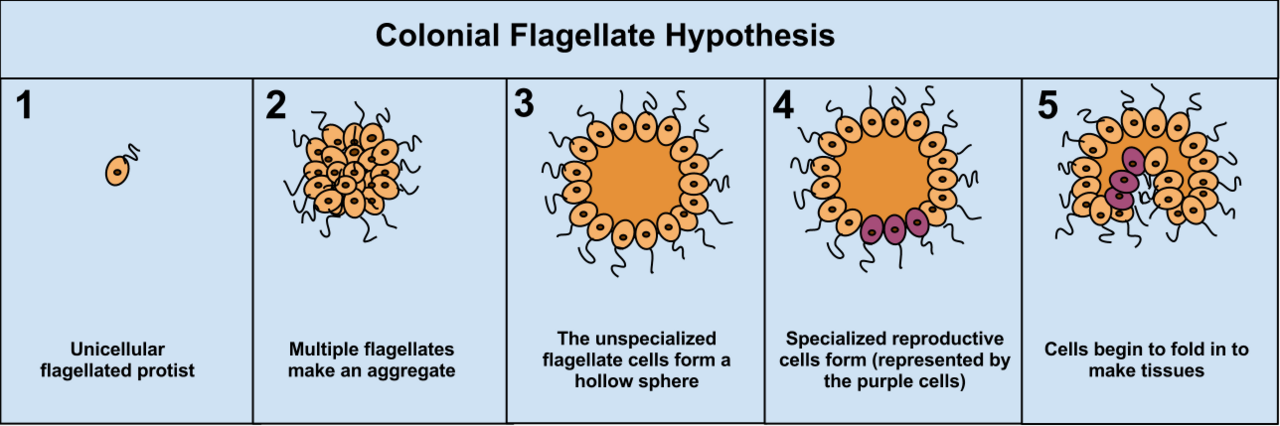
L'avantage de l'hypothèse de la théorie coloniale est qu'elle a été observée indépendamment dans 16 phylums différents du protoctistan. Par exemple, lors de pénuries alimentaires, l'amibe Dictyostelium se regroupe en colonie qui se déplace en une seule vers un nouvel endroit. Certaines de ces amibes se différencient alors légèrement les unes des autres. D'autres exemples d'organisation coloniale chez les protistes sont les Volvocaceae , comme Eudorina et Volvox , ce dernier étant constitué de 500 à 50 000 cellules (selon l'espèce), dont seule une fraction se reproduit. Par exemple, dans une espèce, 25 à 35 cellules se reproduisent, 8 de manière asexuée et environ 15 à 25 de manière sexuée. Cependant, il peut souvent être difficile de séparer les protistes coloniaux des véritables organismes multicellulaires, car les deux concepts ne sont pas distincts ; les protistes coloniaux ont été surnommés « pluricellulaires » plutôt que « multicellulaires ».
La théorie des synzoospores
Certains auteurs suggèrent que l'origine de la multicellularité, au moins chez les métazoaires, s'est produite en raison d'une transition de la différenciation cellulaire temporelle à la différenciation cellulaire spatiale , plutôt que par une évolution progressive de la différenciation cellulaire, comme l'affirme la théorie des gastrées de Haeckel .
GK-PID
Il y a environ 800 millions d'années, un changement génétique mineur dans une seule molécule appelée domaine d'interaction protéique de la guanylate kinase (GK-PID) aurait pu permettre aux organismes de passer d'un organisme unicellulaire à une cellule parmi de nombreuses cellules.
Le rôle des virus
Des gènes empruntés à des virus et des éléments génétiques mobiles (MGE) ont récemment été identifiés comme jouant un rôle crucial dans la différenciation des tissus et organes multicellulaires et même dans la reproduction sexuée, dans la fusion des ovules et des spermatozoïdes. De telles cellules fusionnées sont également impliquées dans les membranes des métazoaires telles que celles qui empêchent les produits chimiques de traverser le placenta et la séparation du corps cérébral. Deux composants viraux ont été identifiés. Le premier est la syncytine , qui provient d'un virus. Le deuxième identifié en 2002 s'appelle EFF-1, qui aide à former la peau de Caenorhabditis elegans , qui fait partie de toute une famille de protéines FF. Felix Rey, de l'Institut Pasteur à Paris, a construit la structure 3D de la protéine EFF-1 et a montré qu'elle fait le travail de liaison d'une cellule à une autre, dans les infections virales. Le fait que toutes les molécules de fusion cellulaire connues soient d'origine virale suggère qu'elles ont joué un rôle vital dans les systèmes de communication intercellulaire qui ont permis la multicellularité. Sans la capacité de fusion cellulaire, des colonies auraient pu se former, mais rien d'aussi complexe qu'une éponge n'aurait été possible.
Hypothèse de la disponibilité de l'oxygène
Cette théorie suggère que l'oxygène disponible dans l'atmosphère de la Terre primitive aurait pu être le facteur limitant de l'émergence de la vie multicellulaire. Cette hypothèse est basée sur la corrélation entre l'émergence de la vie multicellulaire et l'augmentation des niveaux d'oxygène au cours de cette période. Cela aurait eu lieu après la Grande Oxydation mais avant la plus récente augmentation de l'oxygène. Mills conclut que la quantité d'oxygène présente pendant l' Édiacarien n'est pas nécessaire à la vie complexe et qu'il est donc peu probable qu'elle ait été le facteur déterminant de l'origine de la multicellularité.
Hypothèse de la Terre boule de neige
Une Terre boule de neige est un événement géologique au cours duquel toute la surface de la Terre est recouverte de neige et de glace. Le terme peut désigner soit des événements individuels (dont il y en a eu au moins deux), soit une période géologique plus vaste au cours de laquelle toutes les glaciations totales connues se sont produites.
La Terre boule de neige la plus récente a eu lieu pendant la période cryogénique et consistait en deux événements glaciaires mondiaux connus sous le nom de glaciations sturtiennes et marinoennes . Xiao et al . suggèrent qu'entre la période connue sous le nom de « Boring Billion » et la Terre boule de neige, la vie simple aurait pu avoir le temps d'innover et d'évoluer, ce qui pourrait plus tard conduire à l'évolution de la multicellularité.
L'hypothèse de la Terre boule de neige concernant la multicellularité propose que la période cryogénique de l'histoire de la Terre aurait pu être le catalyseur de l'évolution de la vie multicellulaire complexe. Brocks suggère que la période entre le Glacien de Sturtien et le Glacien Marinoen plus récent a permis aux algues planctoniques de dominer les mers, ouvrant la voie à une diversité rapide de la vie pour les lignées végétales et animales. La vie complexe est rapidement apparue et diversifiée dans ce que l'on appelle l' explosion cambrienne peu après le Marinoen.
Hypothèse de prédation
L'hypothèse de la prédation suggère que pour éviter d'être mangés par des prédateurs, les organismes unicellulaires simples ont évolué vers la multicellularité afin de devenir plus difficiles à consommer comme proies. Herron et al. ont réalisé des expériences d'évolution en laboratoire sur l'algue verte unicellulaire, Chlamydomonas reinhardtii , en utilisant la paramécie comme prédateur. Ils ont découvert qu'en présence de ce prédateur, C. reinhardtii développe effectivement des caractéristiques multicellulaires simples.
Evolution expérimentale
Il est impossible de savoir ce qui s’est passé lorsque des cellules individuelles ont évolué en organismes multicellulaires il y a des centaines de millions d’années. Cependant, nous pouvons identifier des mutations qui peuvent transformer des organismes unicellulaires en organismes multicellulaires. Cela démontrerait la possibilité d’un tel événement. Les espèces unicellulaires peuvent acquérir relativement facilement des mutations qui les font s’attacher les unes aux autres – la première étape vers la multicellularité. De nombreuses espèces normalement unicellulaires ont évolué pour présenter ces étapes précoces :
- Les levures sont connues depuis longtemps pour présenter une floculation . L'un des premiers gènes de levure découverts comme étant à l'origine de ce phénotype est FLO1. phénotype plus frappant en amas est appelé « flocon de neige », causé par la perte d'un seul facteur de transcription Ace2 . Les levures « flocon de neige » se développent en amas multicellulaires qui sédimentent rapidement ; elles ont été identifiées par évolution dirigée. Plus récemment (2024), les levures flocon de neige ont été soumises à plus de 3 000 générations d'évolution dirigée supplémentaire, formant des assemblages macroscopiques à l'échelle du millimètre . Des changements dans plusieurs gènes ont été identifiés. En outre, les auteurs ont signalé que seules les cultures anaérobies de levure flocon de neige ont développé ce trait, alors que les cultures aérobies ne l'ont pas fait.
- De nombreuses espèces d'algues vertes ont évolué expérimentalement pour former des touffes plus grandes. Lorsque Chlorella vulgaris est cultivée avec un prédateur Ochromonas vallescia , elle commence à former de petites colonies, qui sont plus difficiles à ingérer en raison de leur plus grande taille. Il en va de même pour Chlamydomonas reinhardtii sous la prédation de Brachionus calyciflorus et Paramecium tetraurelia .
C. reinhartii commence normalement comme une propagule unicellulaire mobile ; cette cellule unique se reproduit de manière asexuée en subissant 2 à 5 cycles de mitose sous forme d'un petit amas de cellules non mobiles, puis toutes les cellules deviennent des propagules unicellulaires et l'amas se dissout. Avec quelques générations sous la prédation de Paramecium , le « groupe » devient une structure persistante : seules quelques cellules deviennent des propagules. Certaines populations vont plus loin et développent des propagules multicellulaires : au lieu de détacher des cellules individuelles de l'amas, l'amas se reproduit maintenant en détachant des amas plus petits.
Avantages
La multicellularité permet à un organisme de dépasser les limites de taille normalement imposées par la diffusion : les cellules individuelles de plus grande taille ont un rapport surface/volume réduit et ont du mal à absorber suffisamment de nutriments et à les transporter dans toute la cellule. Les organismes multicellulaires ont ainsi les avantages compétitifs d'une augmentation de taille sans ses limitations. Ils peuvent avoir une durée de vie plus longue car ils peuvent continuer à vivre lorsque les cellules individuelles meurent. La multicellularité permet également d'accroître la complexité en permettant la différenciation des types de cellules au sein d'un même organisme.
On peut toutefois se demander si tous ces éléments peuvent être considérés comme des avantages : la grande majorité des organismes vivants sont unicellulaires, et même en termes de biomasse, les organismes unicellulaires réussissent bien mieux que les animaux, mais pas que les plantes. Plutôt que de considérer des caractéristiques telles qu'une durée de vie plus longue et une plus grande taille comme un avantage, de nombreux biologistes les voient uniquement comme des exemples de diversité, avec les compromis qui y sont associés.
Modifications de l'expression génétique lors de la transition de l'unicellularité à la multicellularité
Au cours de la transition évolutive des organismes unicellulaires aux organismes multicellulaires, l'expression des gènes associés à la reproduction et à la survie a probablement changé. Dans l'état unicellulaire, les gènes associés à la reproduction et à la survie sont exprimés d'une manière qui améliore la forme physique des cellules individuelles, mais après la transition vers la multicellularité, le modèle d' expression de ces gènes a dû changer de manière substantielle pour que les cellules individuelles deviennent plus spécialisées dans leur fonction relative à la reproduction et à la survie. Au fur et à mesure de l'émergence de l'organisme multicellulaire, les modèles d'expression des gènes se sont compartimentés entre les cellules spécialisées dans la reproduction ( cellules germinales ) et celles spécialisées dans la survie ( cellules somatiques ). Au fur et à mesure de la transition, les cellules spécialisées ont eu tendance à perdre leur propre individualité et ne pouvaient plus à la fois survivre et se reproduire en dehors du contexte du groupe.