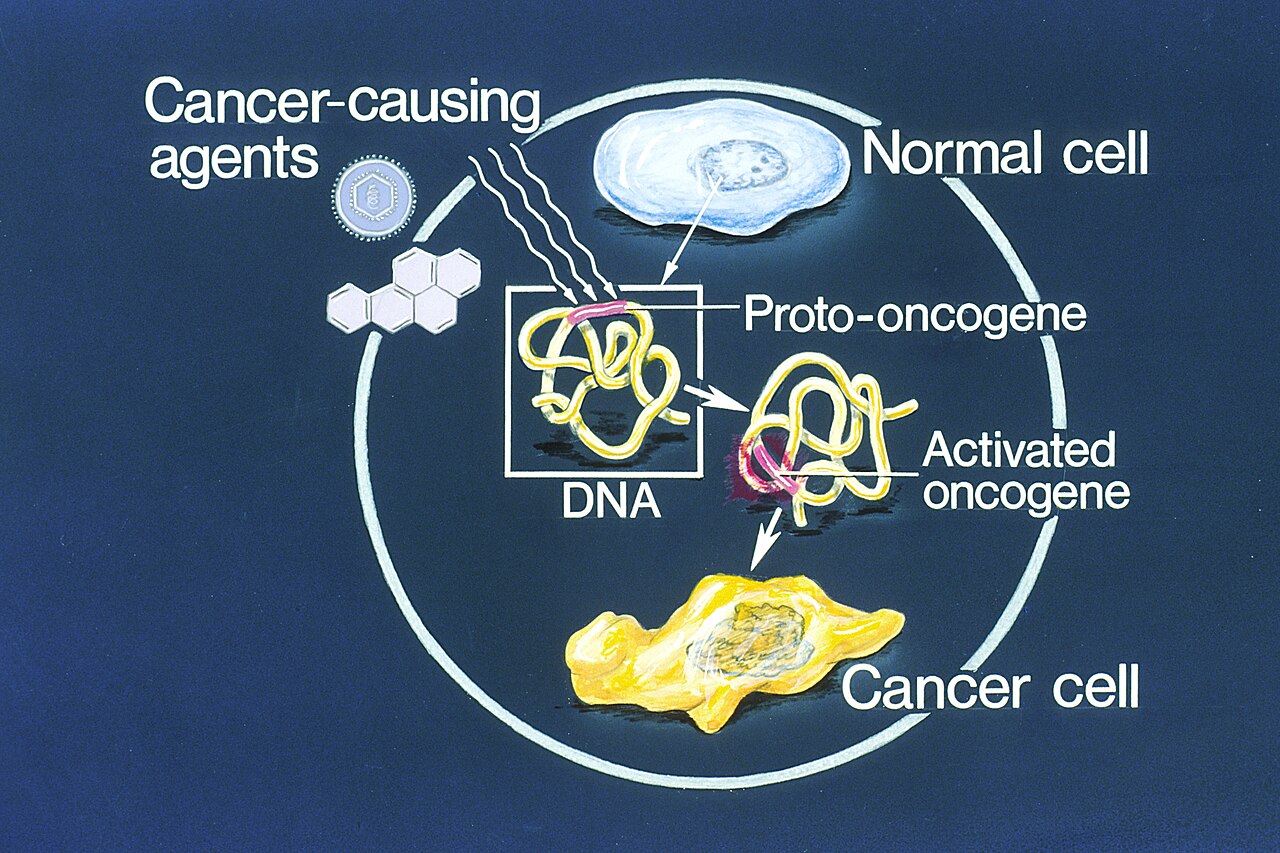
Un oncogène est un gène qui a le potentiel de provoquer un cancer . Dans les cellules tumorales , ces gènes sont souvent mutés ou exprimés à des niveaux élevés.
La plupart des cellules normales subissent une mort cellulaire rapide préprogrammée ( apoptose ) si des fonctions critiques sont altérées et fonctionnent mal. Les oncogènes activés peuvent faire en sorte que les cellules désignées pour l'apoptose survivent et prolifèrent à la place. La plupart des oncogènes ont commencé comme proto-oncogènes : des gènes normaux impliqués dans la croissance et la prolifération cellulaires ou l'inhibition de l'apoptose. Si, par mutation, les gènes normaux favorisant la croissance cellulaire sont régulés à la hausse (mutation à gain de fonction), ils prédisposent la cellule au cancer et sont appelés oncogènes . Habituellement, plusieurs oncogènes, ainsi que des gènes apoptotiques ou suppresseurs de tumeurs mutés , agissent de concert pour provoquer le cancer. Depuis les années 1970, des dizaines d'oncogènes ont été identifiés dans le cancer humain. De nombreux médicaments contre le cancer ciblent les protéines codées par les oncogènes. Les oncogènes sont un ensemble de gènes physiquement et fonctionnellement diversifiés et, par conséquent, leurs produits protéiques ont des effets pléiotropes sur une variété de cascades régulatrices complexes au sein de la cellule.
Les gènes appelés proto-oncogènes sont ceux qui favorisent normalement la croissance et la division cellulaires afin de générer de nouvelles cellules ou de maintenir la viabilité des cellules préexistantes. Lorsqu'ils sont surexprimés, les proto-oncogènes peuvent être activés par inadvertance, ce qui les transforme en oncogènes.
Il existe de nombreuses façons d’activer (de mettre en marche) les oncogènes dans les cellules :
Modifications ou mutations génétiques : le « codage » génétique d'une personne peut différer d'une manière telle qu'un oncogène est toujours activé. Ces types de modifications génétiques peuvent se développer spontanément tout au long de la vie d'une personne ou être héritées d'un parent lorsqu'une erreur de transcription se produit pendant la division cellulaire.
Les cellules peuvent souvent activer ou désactiver des gènes par des mécanismes épigénétiques plutôt que par des altérations génétiques réelles. Par ailleurs, différents composés chimiques pouvant être liés au matériel génétique (ADN ou ARN) peuvent avoir un impact sur les gènes actifs. Un oncogène peut être activé de manière sporadique en raison de ces modifications épigénétiques. Consultez Altérations génétiques et cancer pour en savoir plus sur les altérations épigénétiques.
Réarrangement chromosomique : chaque être vivant possède des chromosomes, qui sont de gros brins d' ADN contenant les gènes d'une cellule. La séquence d'ADN d'un chromosome peut changer à chaque fois qu'une cellule se divise. Cela peut entraîner la localisation d'un gène à proximité d'un proto-oncogène qui agit comme un interrupteur, le gardant actif même lorsqu'il ne devrait pas. La cellule peut se développer de manière irrégulière grâce à ce nouvel oncogène.
Duplication génétique : si une cellule possède plus de copies d’un gène qu’une autre, cette cellule peut produire trop d’une certaine protéine.
Le premier oncogène humain (HRAS), une découverte cruciale dans le domaine de la recherche sur le cancer, a été découvert il y a plus de 40 ans et, depuis, le nombre de nouveaux oncogènes pathogènes n'a cessé d'augmenter. La découverte de petites molécules inhibitrices spécifiques qui ciblent spécifiquement les différentes protéines oncogènes et une analyse mécaniste complète des façons dont les oncogènes dérèglent la signalisation physiologique pour provoquer différents types de cancer et syndromes du développement sont des avancées potentielles futures dans le domaine de la recherche sur le cancer. En étudiant le domaine en pleine expansion de la recherche moléculaire sur les oncogènes, l'objectif de ce numéro spécial était de générer des indicateurs translationnels pratiques qui pourraient répondre aux besoins cliniques.
Les gènes considérés comme essentiels pour le cancer peuvent être divisés en deux catégories selon que les mutations nocives qu'ils contiennent entraînent une perte ou un gain de fonction. Les mutations à gain de fonction des proto-oncogènes poussent les cellules à proliférer alors qu'elles ne devraient pas, tandis que les mutations à perte de fonction des gènes suppresseurs de tumeur libèrent les cellules des inhibitions qui servent généralement à contrôler leur nombre. La capacité des gènes mutants, appelés oncogènes, à orienter une lignée spécifique de cellules tests vers la prolifération maligne peut parfois être utilisée pour identifier ces mutations ultérieures, qui ont un effet dominant.
On a initialement découvert que beaucoup d'entre eux induisaient le cancer chez les animaux lorsqu'ils étaient introduits par une infection à vecteur viral, qui transporte l'information génétique d'une cellule hôte antérieure. Une autre méthode pour identifier les oncogènes consiste à rechercher des gènes activés par des mutations dans les cellules cancéreuses humaines ou par des translocations chromosomiques qui peuvent indiquer la présence d'un gène crucial pour le cancer.
Les patients atteints de cancer sont généralement classés en fonction de paramètres cliniques afin d'adapter leur traitement contre le cancer . Par exemple, la séparation des patients atteints de leucémie aiguë en leucémie lymphocytaire et leucémie myélocytaire est importante, car le traitement optimal pour chaque forme est différent. Même dans une maladie particulière, l'identification des patients ayant un potentiel de pronostic bon et mauvais est utile, car un traitement plus agressif peut être nécessaire pour obtenir une guérison dans le groupe à pronostic mauvais. Les oncogènes sont des marqueurs pronostiques dans certains cancers humains. L'amplification de N-myc est un déterminant indépendant dans la prédiction d'un mauvais pronostic dans le neuroblastome infantile . Les enfants présentant une amplification de N-myc, quel que soit le stade, auront une survie plus courte. Ainsi, les efforts thérapeutiques se concentrent sur l'intensification du traitement dans ce groupe à pronostic mauvais.
Histoire
La théorie des oncogènes a été préfigurée par le biologiste allemand Theodor Boveri dans son livre de 1914 Zur Frage der Entstehung Maligner Tumoren (Concernant l'origine des tumeurs malignes) dans lequel il prédit l'existence d'oncogènes (Teilungsfoerdernde Chromosomen) qui s'amplifient (im permanenten Übergewicht) au cours du développement de la tumeur.
Plus tard, le terme « oncogène » a été redécouvert en 1969 par les scientifiques du National Cancer Institute George Todaro et Robert Huebner .
Le premier oncogène confirmé a été découvert en 1970 et a été appelé SRC (prononcé « sarc » car c'est l'abréviation de sarcome). SRC a été découvert pour la première fois comme oncogène dans un rétrovirus du poulet . Des expériences réalisées par le Dr G. Steve Martin de l' Université de Californie à Berkeley ont démontré que SRC était en effet le gène du virus qui agissait comme oncogène lors de l'infection. La première séquence nucléotidique de v-Src a été séquencée en 1980 par AP Czernilofsky et al.
En 1976, les docteurs Dominique Stéhelin , J. Michael Bishop et Harold E. Varmus de l' Université de Californie à San Francisco ont démontré que les oncogènes étaient des proto-oncogènes activés comme on en trouve dans de nombreux organismes, y compris les humains. Bishop et Varmus ont reçu le prix Nobel de physiologie ou médecine en 1989 pour leur découverte de l'origine cellulaire des oncogènes rétroviraux.
Le Dr Robert Weinberg est reconnu pour avoir découvert le premier oncogène humain identifié dans une lignée cellulaire de cancer de la vessie humaine . La nature moléculaire de la mutation conduisant à l'oncogenèse a ensuite été isolée et caractérisée par le biochimiste espagnol Mariano Barbacid et publiée dans Nature en 1982. allèle muté de HRAS et caractérisant son mécanisme d'activation.
La protéine résultante codée par un oncogène est appelée oncoprotéine . Les oncogènes jouent un rôle important dans la régulation ou la synthèse de protéines liées à la croissance des cellules tumorales. Certaines oncoprotéines sont acceptées et utilisées comme marqueurs tumoraux.
Proto-oncogène
Un proto-oncogène est un gène normal qui pourrait devenir un oncogène en raison de mutations ou d'une expression accrue . Les proto-oncogènes codent pour des protéines qui aident à réguler la croissance et la différenciation cellulaires . Les proto-oncogènes sont souvent impliqués dans la transduction du signal et l'exécution des signaux mitogéniques , généralement par le biais de leurs produits protéiques . Lors de l'acquisition d'une mutation activatrice, un proto-oncogène devient un agent inducteur de tumeur, un oncogène. Les exemples de proto-oncogènes incluent RAS , WNT , MYC , ERK et TRK . Le gène MYC est impliqué dans le lymphome de Burkitt , qui commence lorsqu'une translocation chromosomique déplace une séquence amplificatrice à proximité du gène MYC. Le gène MYC code pour des facteurs de transcription largement utilisés. Lorsque la séquence amplificatrice est mal placée, ces facteurs de transcription sont produits à des taux beaucoup plus élevés. Un autre exemple d'oncogène est le gène Bcr-Abl présent sur le chromosome de Philadelphie , un fragment de matériel génétique observé dans la leucémie myéloïde chronique causée par la translocation de fragments des chromosomes 9 et 22. Bcr-Abl code pour une tyrosine kinase, qui est constitutivement active, conduisant à une prolifération cellulaire incontrôlée. (Plus d'informations sur le chromosome de Philadelphie ci-dessous)
Activation
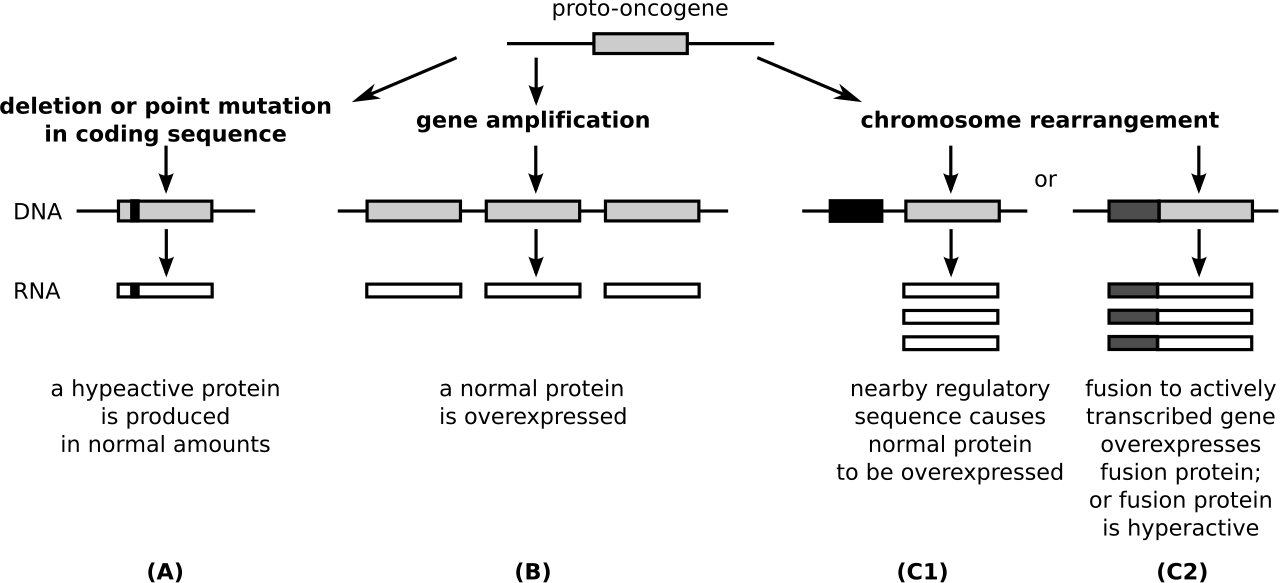
Le proto-oncogène peut devenir un oncogène par une modification relativement minime de sa fonction d'origine. Il existe trois méthodes d'activation de base :
- Une mutation au sein d'un proto-oncogène, ou au sein d'une région régulatrice (par exemple la région promotrice), peut provoquer un changement dans la structure de la protéine, provoquant
- une augmentation de l'activité protéique ( enzymatique )
- une perte de régulation
- Une augmentation de la quantité d’une certaine protéine (concentration en protéines), causée par
- une augmentation de l'expression des protéines (par une mauvaise régulation)
- une augmentation de la stabilité de la protéine (ARNm), prolongeant son existence et donc son activité dans la cellule
- duplication génétique (un type d' anomalie chromosomique ), entraînant une augmentation de la quantité de protéines dans la cellule
- Une translocation chromosomique (un autre type d' anomalie chromosomique )
- Il existe deux types différents de translocations chromosomiques qui peuvent se produire :
- événements de translocation qui déplacent un proto-oncogène vers un nouveau site chromosomique qui conduit à une expression plus élevée
- événements de translocation qui conduisent à une fusion entre un proto-oncogène et un 2e gène (cela crée une protéine de fusion avec une activité cancéreuse/oncogène accrue)
- l'expression d'une protéine hybride constitutivement active . Ce type de mutation dans une cellule souche en division dans la moelle osseuse conduit à la leucémie de l'adulte
- Le chromosome de Philadelphie est un exemple de ce type d'événement de translocation. Ce chromosome a été découvert en 1960 par Peter Nowell et David Hungerford, et il s'agit d'une fusion de parties d'ADN du chromosome 22 et du chromosome 9. L'extrémité cassée du chromosome 22 contient le gène « BCR », qui fusionne avec un fragment du chromosome 9 qui contient le gène « ABL1 ». Lorsque ces deux fragments de chromosome fusionnent, les gènes fusionnent également, créant un nouveau gène : « BCR-ABL ». Ce gène fusionné code pour une protéine qui présente une activité de protéine tyrosine kinase élevée (cette activité est due à la moitié « ABL1 » de la protéine). L'expression non régulée de cette protéine active d'autres protéines impliquées dans le cycle cellulaire et la division cellulaire, ce qui peut provoquer une croissance et une division incontrôlables d'une cellule (la cellule devient cancéreuse). En conséquence, le chromosome de Philadelphie est associé à la leucémie myéloïde chronique (comme mentionné précédemment) ainsi qu'à d'autres formes de leucémie.
L'expression des oncogènes peut être régulée par des microARN (miARN), de petits ARN de 21 à 25 nucléotides de longueur qui contrôlent l'expression des gènes en les régulant à la baisse . Les mutations dans ces microARN (appelés oncomirs ) peuvent conduire à l'activation des oncogènes. antisens pourraient théoriquement être utilisés pour bloquer les effets des oncogènes.
Classification
Il existe plusieurs systèmes de classification des oncogènes, mais il n'existe pas encore de norme largement acceptée. Ils sont parfois regroupés à la fois spatialement (de l'extérieur de la cellule vers l'intérieur) et chronologiquement (parallèlement au processus « normal » de transduction du signal). Plusieurs catégories sont couramment utilisées :
Les propriétés supplémentaires du régulateur oncogénétique comprennent :
- Les facteurs de croissance sont généralement sécrétés par des cellules spécialisées ou non spécialisées pour induire une prolifération cellulaire dans elles-mêmes, dans des cellules voisines ou à distance. Un oncogène peut amener une cellule à sécréter des facteurs de croissance même si elle ne le fait pas normalement. Il induira ainsi sa propre prolifération incontrôlée ( boucle autocrine ) et la prolifération des cellules voisines, ce qui pourrait conduire à la formation de tumeurs. Il peut également provoquer la production d'hormones de croissance dans d'autres parties du corps.
- Les récepteurs tyrosine kinases ajoutent des groupes phosphate à d'autres protéines afin de les activer ou de les désactiver. Les récepteurs kinases ajoutent des groupes phosphate aux protéines réceptrices à la surface de la cellule (qui reçoivent des signaux protéiques de l'extérieur de la cellule et les transmettent à l'intérieur de la cellule). Les tyrosine kinases ajoutent des groupes phosphate à l'acide aminé tyrosine dans la protéine cible. Elles peuvent provoquer le cancer en activant le récepteur de manière permanente (de manière constitutive), même sans signaux provenant de l'extérieur de la cellule.
- Ras est une petite GTPase qui hydrolyse le GTP en GDP et en phosphate. Ras est activée par la signalisation des facteurs de croissance (c.-à-d. EGF, TGFbeta) et agit comme un interrupteur binaire (marche/arrêt) dans les voies de signalisation de croissance. Les effecteurs en aval de Ras comprennent trois protéines kinases activées par les mitogènes Raf a MAP Kinase Kinase Kinase (MAPKKK), MEK a MAP Kinase Kinase (MAPKK) et ERK a MAP Kinase (MAPK), qui à leur tour régulent les gènes qui interviennent dans la prolifération cellulaire.