La pyocyanine (PCN − ) est l'un des nombreux composés toxiques produits et sécrétés par la bactérie Gram négative Pseudomonas aeruginosa . La pyocyanine est un métabolite secondaire bleu, devenant rouge en dessous de pH 4,9, avec la capacité d'oxyder et de réduire d'autres molécules et donc de tuer les microbes en compétition avec P. aeruginosa ainsi que les cellules pulmonaires de mammifères que P. aeruginosa a infectées au cours de la mucoviscidose . La pyocyanine étant un zwitterion au pH sanguin, elle est facilement capable de traverser la membrane cellulaire. La pyocyanine peut exister dans trois états différents : oxydée (bleue), réduite de manière monovalente (incolore) ou réduite de manière divalente (rouge). Les mitochondries jouent un rôle important dans le cycle de la pyocyanine entre ses états redox. En raison de ses propriétés redox-actives, la pyocyanine génère des espèces réactives de l'oxygène .
Biosynthèse
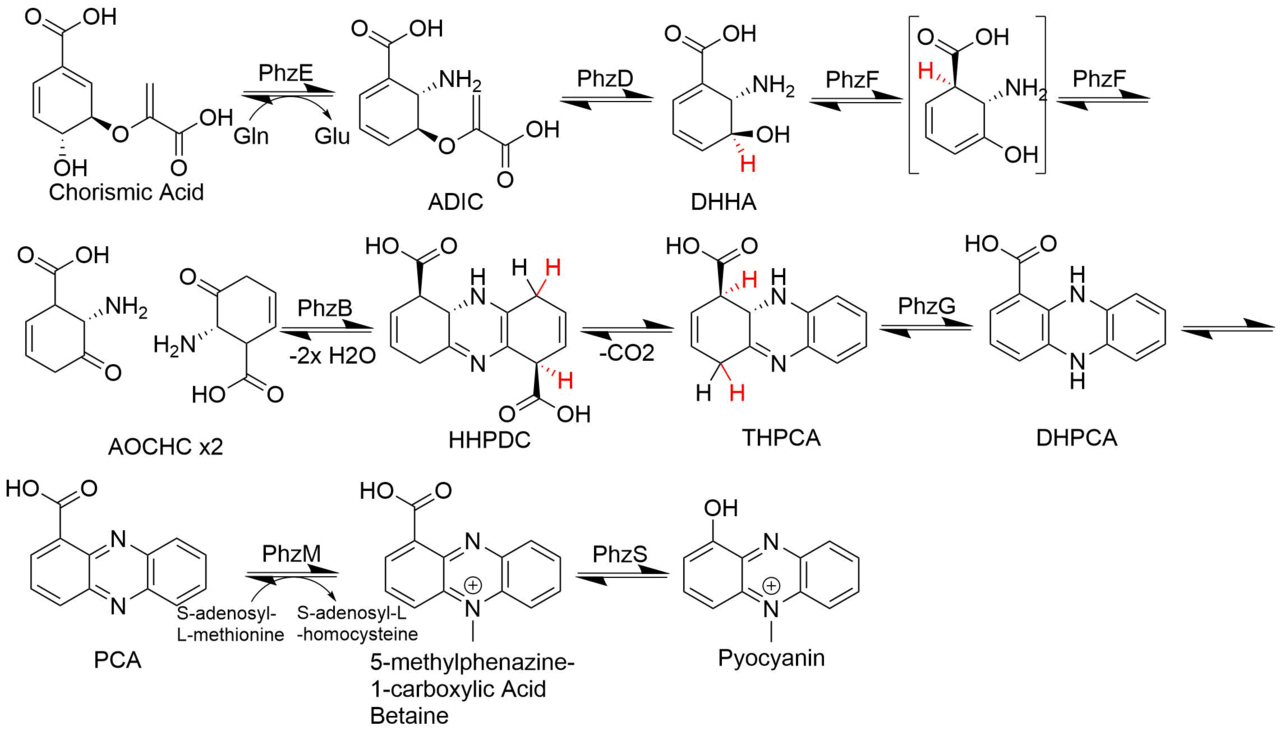
La biosynthèse de la pyocyanine commence par la synthèse du noyau de l'acide phénazine-1-carboxylique (PCA). Dans cette réaction, l'enzyme PhzE catalyse la perte du groupe hydroxyle du C4 de l'acide chorismique ainsi que le transfert d'un groupe amine de la glutamine pour former l'acide glutamique et l'acide 2-amino-2-désoxyisochorismique (ADIC). Ensuite, PhzD catalyse l'élimination hydrolytique de la fraction pyruvate de l'ADIC pour former l'acide (5S,6S)-6-amino-5-hydroxy-1,3-cyclohexadiève-1-carboxylique (DHHA). Dans l'étape suivante, PhzF catalyse deux étapes : l'abstraction d'un hydrogène du C3 du DHHA, la délocalisation du système de double liaison et la reprotonation en C1 ainsi que la tautomérisation de l'énol pour former l'acide 6-amino-5-oxocyclohex-2-ène-1-carboxylique (AOCHC) hautement instable. À partir de là, deux molécules d'AOCHC sont condensées par PhzB pour former le composé tricyclique, l'acide hexahydrophénazine-1,6-dicarboxylique (HHPDC). Le produit de cette réaction, HHPDC, est instable et subit spontanément une décarboxylation oxydative dans une réaction non catalysée pour former l'acide tétrahydrophénazine-1,6-carboxylique (THPCA). Dans l'étape finale de la synthèse de l'acide phénazine-1-carboxylique, l'enzyme PhzG catalyse l'oxydation du THPCA en acide dihydro-phénazine-1-carboxylique. Il s'agit de la dernière étape catalysée dans la production de PCA, la dernière étape est une oxydation non catalysée de DHPCA en PCA. La conversion du PCA en pyocyanine est réalisée en deux étapes enzymatiques : tout d'abord, le PCA est méthylé sur N5 en 5-méthylphénazine-1-carboxylate bétaïne par l'enzyme PhzM en utilisant le cofacteur S-adénosyl-L-méthionine et deuxièmement, PhzS catalyse la décarboxylation hydroxylative de ce substrat pour former le produit final, la pyocyanine.
Les chromosomes de la plupart des souches de P. aeruginosa portent deux opérons presque identiques, phzA1B1C1D1E1F1G1 et phzA2B2C2D2E2F2G2 , qui codent les enzymes nécessaires à la production de PCA. La transcription de ces opérons est contrôlée par la détection de quorum , et plus spécifiquement par le système Pseudomonas Quinolone Signal (PQS) impliquant le régulateur transcriptionnel MvfR (également connu sous le nom de PqsR). La conversion de PCA en pyocyanine est ensuite réalisée par les produits de phzS et phzS , qui sont des gènes uniques dans le chromosome. La biosynthèse peut être altérée en perturbant la voie aro qui est nécessaire à la synthèse de l'acide chorismique à partir du shikimate.
Guerre redox
La pyocyanine inactive la catalase en réduisant la transcription de son gène et en ciblant directement l'enzyme elle-même. Le glutathion est un antioxydant important modulé par la pyocyanine. En particulier, le pool de la forme réduite est épuisé tandis que la forme oxydée est favorisée par le peroxyde d'hydrogène qui n'est pas dismuté par la catalase. Dans le poumon atteint de mucoviscidose, la pyocyanine intracellulaire convertit l'oxygène moléculaire en radical libre superoxyde en oxydant le NADPH en NADP + . Cela a un double effet négatif sur les poumons. Tout d'abord, le NADPH utilisé par la pyocyanine épuise le substrat disponible pour la réaction catalysée par l' enzyme NADPH oxydase . Deuxièmement, le radical superoxyde généré peut inhiber les cytokines , telles que l'IL-4 , l'IL-13 et l'IFN-γ , qui régulent généralement à la hausse la NADPH oxydase. Lorsque le poumon est confronté à la pyocyanine, une concentration accrue de catalase et de superoxyde dismutase est observée afin de faire face au barrage de radicaux produits.
Objectifs
La pyocyanine est capable de cibler un large éventail de composants et de voies cellulaires. Les voies affectées par la pyocyanine comprennent la chaîne de transport d'électrons , le transport vésiculaire et la croissance cellulaire. Une sensibilité accrue à la pyocyanine est observée dans les cellules contenant certaines protéines ou certains complexes mutants. Les mutations des gènes affectant la synthèse et l'assemblage de la V-ATPase , la machinerie de transport des vésicules et la machinerie de tri des protéines confèrent toutes une sensibilité accrue à la pyocyanine, ce qui renforce encore les effets sur la mucoviscidose chez le patient. L'ATPase vacuolaire dans les cellules de levure est une cible particulièrement puissante car elle est le principal producteur non mitochondrial d'ATP, mais elle a également de nombreuses autres fonctions telles que le contrôle homéostatique du calcium, la facilitation de l'endocytose médiée par les récepteurs et la dégradation des protéines. Par conséquent, l'inactivation de l'ATPase vacuolaire par le peroxyde d'hydrogène produit par la pyocyanine a d'énormes conséquences pour les poumons. Outre ces effets, la pyocyanine cible également les protéases de type caspase 3 qui peuvent ensuite déclencher l'apoptose et la nécrose . Les transporteurs d'électrons mitochondriaux ubiquinone et acide nicotinique sont également sensibles à la pyocyanine. Le cycle cellulaire peut être perturbé par l'action de la pyocyanine, et elle peut entraver la prolifération des lymphocytes . Cela se produit par la génération d' intermédiaires réactifs de l'oxygène , tels que le peroxyde d'hydrogène et le superoxyde , qui provoquent un stress oxydatif en endommageant directement l'ADN ou en ciblant d'autres constituants du cycle cellulaire tels que la recombinaison de l'ADN et le mécanisme de réparation. La pyocyanine contribue à la disproportion de l'activité protéase et antiprotéase en désactivant l'inhibiteur de la protéase α 1 .
Fibrose kystique
De nombreuses études ont conclu que la pyocyanine a un effet néfaste sur la fibrose kystique, ce qui permet à P. aeruginosa de persister dans les poumons atteints de fibrose kystique ; elle est souvent détectée dans les expectorations des patients atteints de fibrose kystique. La pyocyanine in vitro a la capacité d'interférer avec des fonctions telles que le battement ciliaire et donc de provoquer un dysfonctionnement épithélial, car les cils sont nécessaires pour balayer le mucus dans la gorge. De plus, l'apoptose des neutrophiles , la libération d'immunoglobulines par les lymphocytes B et la libération d'interleukines (par exemple IL-8 et CCL5 ) sont toutes altérées par la pyocyanine, affaiblissant le système immunitaire du poumon. Des études in vivo ont montré que la croissance du champignon est inhibée en présence de pyocyanine. Le mécanisme fongicide est l'activation du NAD(P)H pour induire une cascade redox-active produisant des intermédiaires réactifs de l'oxygène. Cela permet à P. aeruginosa d'avoir un avantage compétitif car il peut dominer les autres micro-organismes dans le poumon atteint de mucoviscidose. La concentration intracellulaire d' ATP est également diminuée par la pyocyanine, ce qui provoque des dommages supplémentaires au CFTR qui sont déjà altérés dans la mucoviscidose. Les canaux CFTR dépendent de l'ATP pour deux raisons principales. Tout d'abord, la liaison et l'hydrolyse de l'ATP doivent se produire au niveau de deux domaines de liaison nucléotidique pour que le canal passe de sa conformation ouverte à sa conformation fermée. Deuxièmement, la phosphorylation du CFTR par la protéine kinase A doit se produire pour que le canal soit opérationnel. La PKA est activée par l'AMPc qui est produit à partir de l'ATP. Ces deux processus sont altérés lorsque l'ATP est épuisé par la pyocyanine.
Défense contre la pyocyanine
Caenorhabditis elegans possède deux transporteurs ABC spécifiques appelés pgp-1 et pgp-2 qui sont capables d'extruder efficacement la pyocyanine intracellulaire de manière dépendante de l'énergie.