Les petits ARN nucléaires ( snRNA ) sont une classe de petites molécules d'ARN que l'on trouve dans les taches d'épissage et les corps de Cajal du noyau cellulaire des cellules eucaryotes . La longueur moyenne d'un snRNA est d'environ 150 nucléotides. Ils sont transcrits par l'ARN polymérase II ou l'ARN polymérase III . Leur fonction principale est le traitement de l'ARN pré-messager ( hnRNA ) dans le noyau. Il a également été démontré qu'ils contribuent à la régulation des facteurs de transcription ( ARN 7SK ) ou de l'ARN polymérase II (ARN B2) et au maintien des télomères .
Les snRNA sont toujours associés à un ensemble de protéines spécifiques, et les complexes sont appelés petites ribonucléoprotéines nucléaires ( snRNP , souvent prononcées « snurps »). Chaque particule snRNP est composée d'un composant snRNA et de plusieurs protéines spécifiques aux snRNP (y compris les protéines Sm , une famille de protéines nucléaires). Les composants snRNA humains les plus courants de ces complexes sont connus, respectivement, sous les noms suivants : ARN épissosomal U1 , ARN épissosomal U2 , ARN épissosomal U4 , ARN épissosomal U5 et ARN épissosomal U6 . Leur nomenclature dérive de leur teneur élevée en uridine .
Les snRNA ont été découverts par accident lors d'une expérience d'électrophorèse sur gel en 1966. Un type d'ARN inattendu a été trouvé dans le gel et étudié. Des analyses ultérieures ont montré que ces ARN étaient riches en uridylate et s'établissaient dans le noyau.
Les snRNA et les petits ARN nucléolaires (snoRNA) ne sont pas identiques et aucun des deux n'est un sous-type de l'autre. Tous deux sont différents et appartiennent à une classe de petits ARN. Il s'agit de petites molécules d'ARN qui jouent un rôle essentiel dans la biogenèse de l'ARN et guident les modifications chimiques des ARN ribosomiques (ARNr) et d'autres gènes d'ARN (ARNt et snRNA). Ils sont situés dans le nucléole et les corps de Cajal des cellules eucaryotes (les principaux sites de synthèse de l'ARN), où ils sont appelés scaRNA (petits ARN spécifiques du corps de Cajal).
Cours
Les snRNA sont souvent divisés en deux classes en fonction des caractéristiques de séquence communes ainsi que des facteurs protéiques associés tels que les protéines LSm de liaison à l'ARN .
La première classe, connue sous le nom de snRNA de classe Sm , est plus largement étudiée et se compose de U1, U2, U4, U4atac , U5, U7 , U11 et U12 . Les snRNA de classe Sm sont transcrits par l'ARN polymérase II . Les pré-snRNA sont transcrits et reçoivent la coiffe habituelle de 7-méthylguanosine à cinq primes dans le noyau . Ils sont ensuite exportés vers le cytoplasme par les pores nucléaires pour un traitement ultérieur. Dans le cytoplasme, les snRNA reçoivent une coupe 3' pour former une structure tige-boucle 3', ainsi qu'une hyperméthylation de la coiffe 5' pour former la triméthylguanosine. La structure tige 3' est nécessaire à la reconnaissance par la protéine de survie du motoneurone (SMN). Ce complexe assemble les snRNA en ribonucléoprotéines stables (RNP). La coiffe 5' modifiée est alors nécessaire pour réimporter le snRNP dans le noyau. Tous ces snRNA riches en uridine, à l'exception de U7, forment le noyau du spliceosome . L'épissage, ou la suppression des introns , est un aspect majeur de la modification post-transcriptionnelle et n'a lieu que dans le noyau des eucaryotes. Il a été découvert que le snRNA U7 fonctionne dans le traitement du pré-ARNm des histones .
La deuxième classe, connue sous le nom de snRNA de classe Lsm , se compose de U6 et U6atac . Les snRNA de classe Lsm sont transcrits par l'ARN polymérase III et ne quittent jamais le noyau, contrairement aux snRNA de classe Sm. Les snRNA de classe Lsm contiennent une coiffe 5'-γ-monométhylphosphate et une tige-boucle 3', se terminant par un segment d'uridines qui forment le site de liaison d'un anneau hétéroheptamérique distinct de protéines Lsm.
Dans le spliceosome
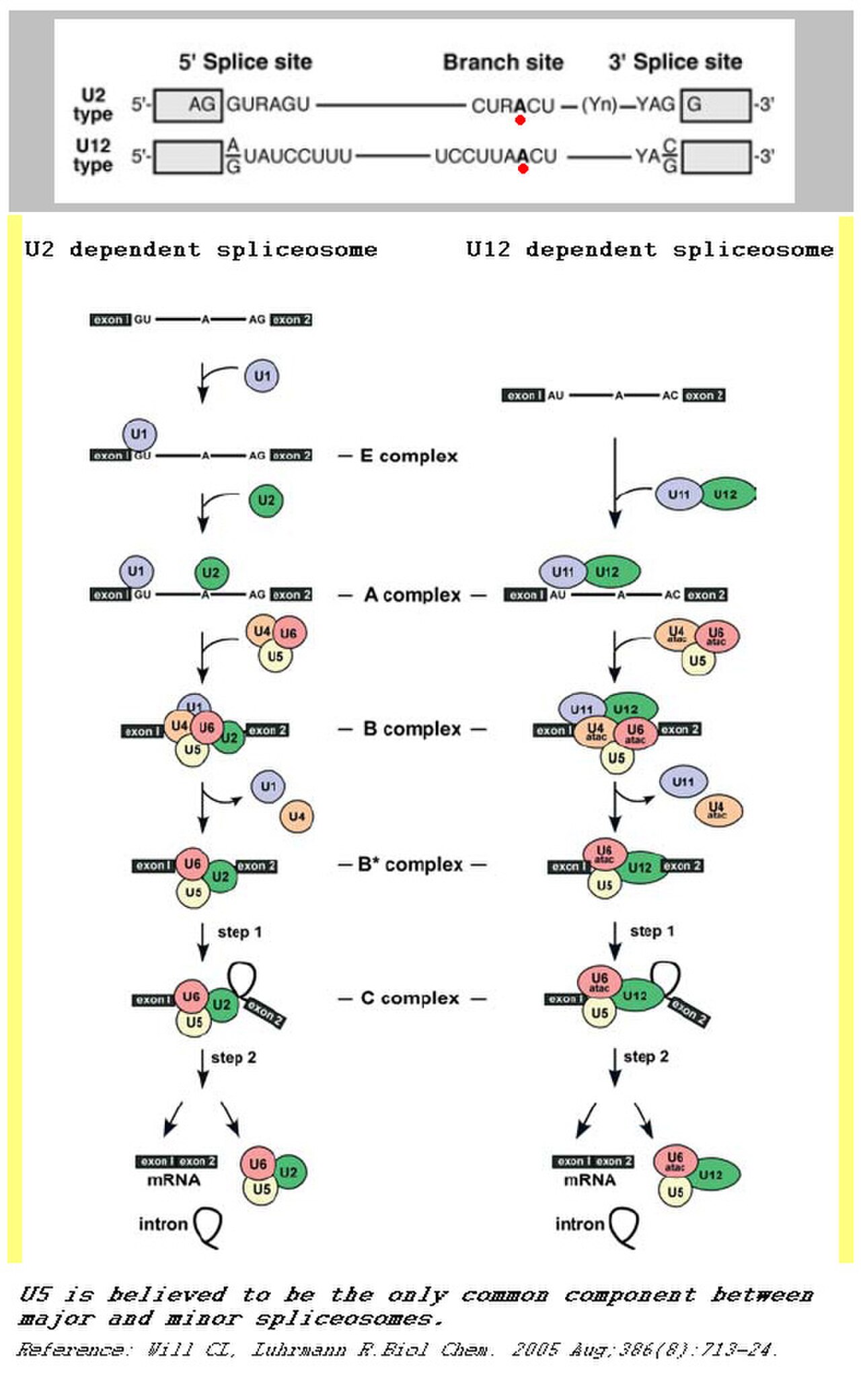
Les spliceosomes catalysent l'épissage , une étape essentielle de la maturation de l'ARN messager précurseur eucaryote. Une erreur d'épissage même dans un seul nucléotide peut être dévastatrice pour la cellule, et une méthode fiable et répétable de traitement de l'ARN est nécessaire pour assurer la survie cellulaire. Le spliceosome est un grand complexe protéine-ARN composé de cinq petits ARN nucléaires (U1, U2, U4, U5 et U6) et de plus de 150 protéines. Les snRNA, ainsi que leurs protéines associées, forment des complexes ribonucléoprotéiques (snRNP), qui se lient à des séquences spécifiques sur le substrat pré-ARNm . Ce processus complexe entraîne deux réactions de transestérification séquentielles. Ces réactions produiront un intron lariat libre et lieront deux exons pour former un ARNm mature. Il existe deux classes distinctes de spliceosomes. La classe principale, qui est beaucoup plus abondante dans les cellules eucaryotes, épisse principalement les introns de type U2. L'étape initiale de l'épissage est la liaison du snRNP U1 et de ses protéines associées à l'extrémité d'épissure 5' de l' ARNhn . Cela crée le complexe d'engagement qui contraindra l'ARNhn à la voie d'épissage. Ensuite, le snRNP U2 est recruté sur le site de liaison du spliceosome et forme le complexe A, après quoi le complexe tri-snRNP U5.U4/U6 se lie au complexe A pour former la structure connue sous le nom de complexe B. Après réarrangement, le complexe C est formé et le spliceosome est actif pour la catalyse. Dans le spliceosome catalytiquement actif, les snARN U2 et U6 se replient pour former une structure conservée appelée triplex catalytique. Cette structure coordonne deux ions magnésium qui forment le site actif du spliceosome. Il s'agit d'un exemple de catalyse d'ARN .
En plus de ce complexe épissosome principal, il existe un épissosome mineur beaucoup moins courant (~1 %) . Ce complexe comprend les snRNP U11, U12, U4atac, U6atac et U5. Ces snRNP sont des analogues fonctionnels des snRNP utilisés dans l'épissosome majeur. L'épissosome mineur épisse les introns de type U12. Les deux types d'introns diffèrent principalement par leurs sites d'épissage : les introns de type U2 ont des sites d'épissage GT-AG 5' et 3' tandis que les introns de type U12 ont des sites d'épissage AT-AC à leurs extrémités 5' et 3'. L'épissosome mineur exerce sa fonction par une voie différente de l'épissosome majeur.
ARNsn U1
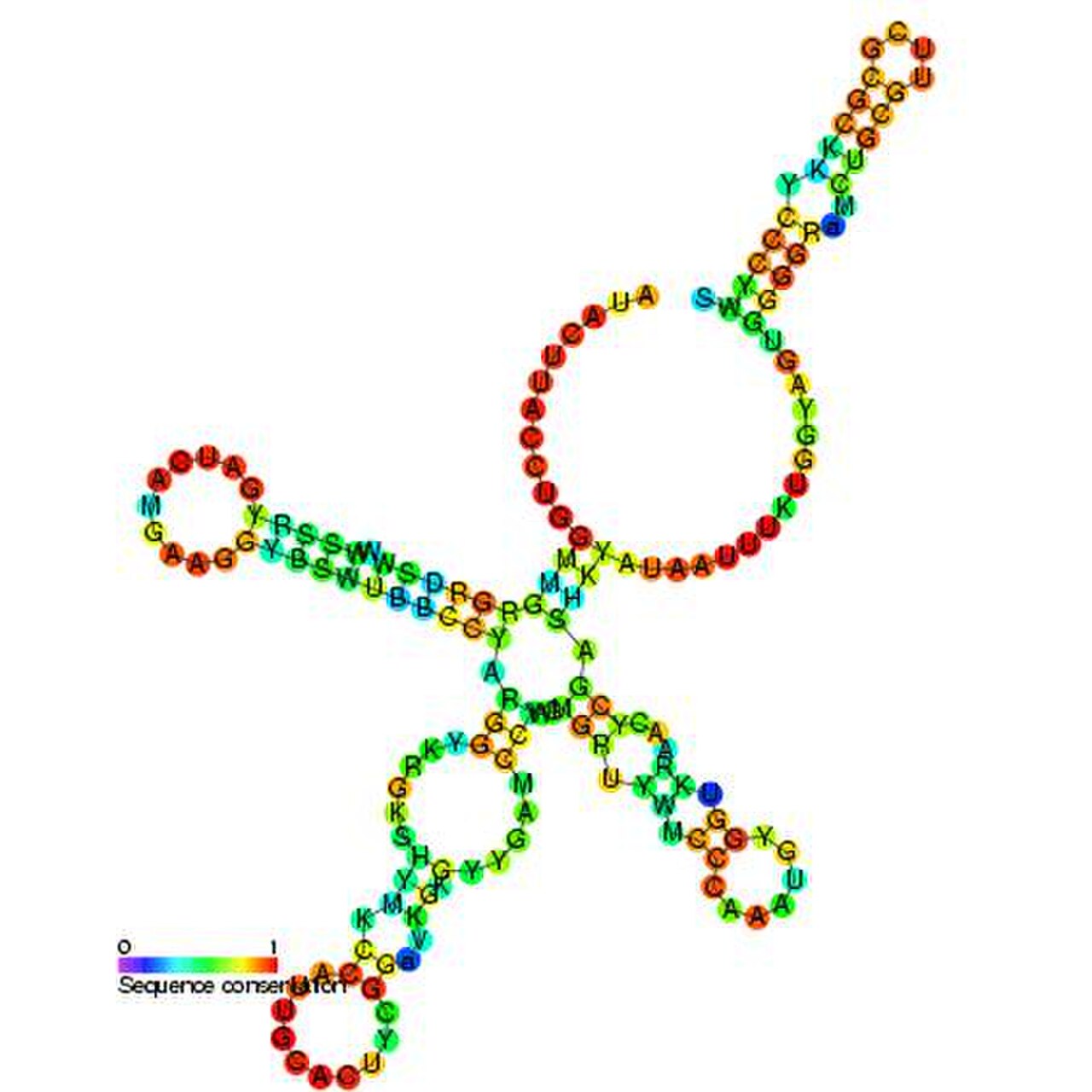
L'U1 snRNP est l'initiateur de l'activité spliceosomale dans la cellule par appariement de bases avec le site d'épissage 5' du pré-ARNm. Dans le spliceosome majeur, des données expérimentales ont montré que l'U1 snRNP est présent en stoechiométrie égale avec les U2, U4, U5 et U6 snRNP. Cependant, l'abondance de l'U1 snRNP dans les cellules humaines est bien supérieure à celle des autres snRNP. Grâce à l'inactivation du gène U1 snRNA dans les cellules HeLa , des études ont montré que l'U1 snRNA revêt une grande importance pour la fonction cellulaire. Lorsque les gènes U1 snRNA ont été inactivés, les microarrays génomiques ont montré une accumulation accrue de pré-ARNm non épissé. De plus, il a été démontré que l'inactivation provoquait un clivage prématuré et une polyadénylation principalement dans les introns situés près du début de la transcription. Lorsque d'autres snRNA à base d'uridine ont été éliminés, cet effet n'a pas été observé. Ainsi, il a été démontré que l'appariement de bases snRNA U1–pré-ARNm protège le pré-ARNm de la polyadénylation ainsi que du clivage prématuré. Cette protection spéciale peut expliquer la surabondance de snRNA U1 dans la cellule.
Les snRNP et les maladies humaines
Grâce à l’étude des petites ribonucléoprotéines nucléaires (snRNP) et des petites RNP nucléolaires (sno), nous avons pu mieux comprendre de nombreuses maladies importantes.
Atrophie musculaire spinale - Les mutations du gène du motoneurone de survie 1 (SMN1) entraînent la dégénérescence des motoneurones spinaux et une atrophie musculaire sévère. La protéine SMN assemble les snRNP de classe Sm, et probablement aussi les snoRNP et d'autres RNP. L'atrophie musculaire spinale touche jusqu'à 1 personne sur 6 000 et constitue la deuxième cause de maladie neuromusculaire , après la dystrophie musculaire de Duchenne .
Dyskératose congénitale – Des mutations dans les snRNP assemblés sont également à l’origine de la dyskératose congénitale, un syndrome rare qui se manifeste par des modifications anormales de la peau, des ongles et des muqueuses. Parmi les effets ultimes de cette maladie figurent l’insuffisance de la moelle osseuse ainsi que le cancer. Il a été démontré que ce syndrome résulte de mutations dans plusieurs gènes, notamment la dyskérine , l’ARN télomérase et la transcriptase inverse de la télomérase .
Syndrome de Prader-Willi – Ce syndrome touche jusqu’à 1 personne sur 12 000 et se manifeste par une faim extrême, des problèmes cognitifs et comportementaux, un faible tonus musculaire et une petite taille. Le syndrome a été lié à la suppression d’une région du chromosome paternel 15 qui n’est pas exprimée sur le chromosome maternel. Cette région comprend un snRNA spécifique du cerveau qui cible l’ ARNm du récepteur de la sérotonine -2C.
Médulloblastome – L’ARNsn U1 est muté dans un sous-ensemble de ces tumeurs cérébrales et conduit à un épissage altéré de l’ARN . Les mutations surviennent principalement dans les tumeurs adultes et sont associées à un mauvais pronostic.
Modification post-transcriptionnelle
Chez les eucaryotes , les snRNA contiennent une quantité significative de modifications de méthylation 2'-O et de pseudouridylations . Ces modifications sont associées à l'activité des snoRNA qui modifient canoniquement les ARNr prématurés, mais ont été observées dans la modification d'autres cibles ARN cellulaires telles que les snRNA. Enfin, l'oligo-adénylation (courte queue poly(A)) peut déterminer le sort des snRNA (qui ne sont généralement pas à queue poly(A)) et ainsi induire leur dégradation en ARN. Ce mécanisme régulant l'abondance des snRNA est à son tour couplé à un changement généralisé de l'épissage alternatif de l'ARN.