En biologie moléculaire , les petits ARN nucléolaires ( snoRNA ) sont une classe de petites molécules d'ARN qui guident principalement les modifications chimiques d'autres ARN, principalement les ARN ribosomiques , les ARN de transfert et les petits ARN nucléaires . Il existe deux classes principales de snoRNA, les snoRNA à boîte C/D, qui sont associés à la méthylation , et les snoRNA à boîte H/ACA, qui sont associés à la pseudouridylation . Les snoRNA sont communément appelés ARN guides, mais ne doivent pas être confondus avec les ARN guides qui dirigent l'édition d'ARN dans les trypanosomes ou les ARN guides (gRNA) utilisés par Cas9 pour l'édition de gènes CRISPR .
Modifications guidées par snoRNA
Après la transcription , les molécules d'ARNr naissantes (appelées pré-ARNr) subissent une série d'étapes de traitement pour générer la molécule d'ARNr mature. Avant le clivage par les exo- et endonucléases, le pré-ARNr subit un schéma complexe de modifications nucléosidiques. Il s'agit notamment de méthylations et de pseudouridylations, guidées par des snoARN.
- La méthylation est la fixation ou la substitution d'un groupe méthyle sur divers substrats . L'ARNr des humains contient environ 115 modifications du groupe méthyle. La majorité d'entre elles sont des méthylations 2'O-ribose (où le groupe méthyle est fixé au groupe ribose).
- La pseudo-uridylation est la conversion ( isomérisation ) du nucléoside uridine en une forme isomérique différente, la pseudo-uridine (Ψ). Cette modification consiste en une rotation de 180º de la base uridine autour de sa liaison glycosyle au ribose du squelette de l'ARN. Après cette rotation, la base azotée apporte un atome de carbone à la liaison glycosyle au lieu de l'atome d'azote habituel. L'aspect bénéfique de cette modification est le donneur de liaison hydrogène supplémentaire disponible sur la base. Alors que l'uridine crée deux liaisons hydrogène avec sa paire de bases Watson-Crick, l'adénine, la pseudo-uridine est capable de créer trois liaisons hydrogène. Lorsque la pseudo-uridine est appariée à l'adénine, elle peut également créer une autre liaison hydrogène, ce qui permet à la complexité de la structure de l'ARNr mature de prendre forme. Le donneur de liaison hydrogène libre forme souvent une liaison avec une base éloignée d'elle-même, créant la structure tertiaire que l'ARNr doit avoir pour être fonctionnel. Les ARNr humains matures contiennent environ 95 modifications Ψ.
snoRNP
Chaque molécule de snoRNA agit comme un guide pour une (ou deux) seule(s) modification(s) individuelle(s) dans un ARN cible. Afin de réaliser la modification, chaque snoRNA s'associe à au moins quatre protéines de base dans un complexe ARN/protéine appelé petite particule ribonucléoprotéique nucléolaire (snoRNP). Les protéines associées à chaque ARN dépendent du type de molécule de snoRNA (voir les familles de guides de snoRNA ci-dessous). La molécule de snoRNA contient un élément antisens (un tronçon de 10 à 20 nucléotides ), qui est complémentaire de la base de la séquence entourant la base ( nucléotide ) ciblée pour la modification dans la molécule de pré-ARN. Cela permet au snoRNP de reconnaître et de se lier à l'ARN cible. Une fois que le snoRNP s'est lié au site cible, les protéines associées sont à l'emplacement physique correct pour catalyser la modification chimique de la base cible.
familles guides snoRNA
Les deux types différents de modification de l'ARNr (méthylation et pseudouridylation) sont dirigés par deux familles différentes de snoRNA. Ces familles de snoRNA sont appelées snoRNA antisens C/D box et H/ACA box en raison de la présence de motifs de séquence conservés dans le snoRNA. Il existe des exceptions, mais en règle générale, les membres de la boîte C/D guident la méthylation et les membres H/ACA guident la pseudouridylation. Les membres de chaque famille peuvent varier en termes de biogenèse, de structure et de fonction, mais chaque famille est classée selon les caractéristiques généralisées suivantes. Pour plus de détails, voir la revue. Les snoRNA sont classés sous la catégorie des petits ARN nucléaires dans MeSH . Le HGNC , en collaboration avec snoRNABase et des experts du domaine, a approuvé des noms uniques pour les gènes humains qui codent les snoRNA.
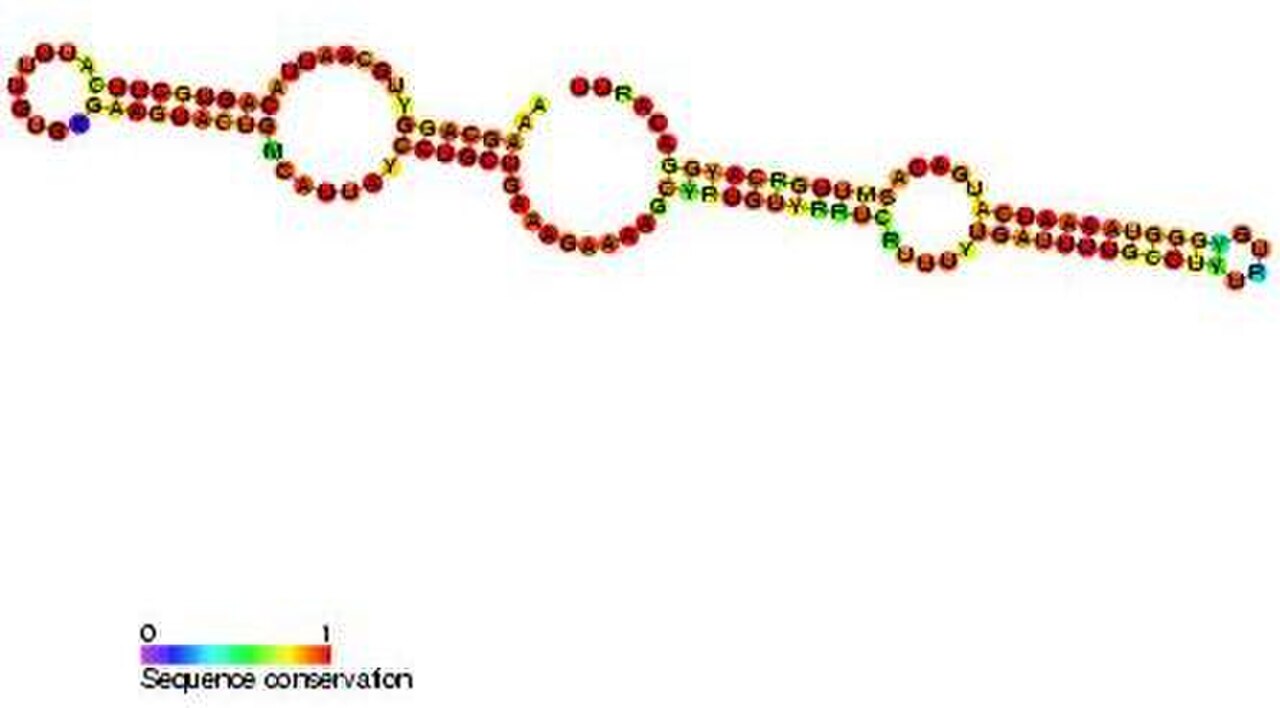
Boîte C/D
Les snoRNA à boîte C/D contiennent deux courts motifs de séquence conservés, C (RUGAUGA) et D (CUGA), situés respectivement près des extrémités 5' et 3' du snoRNA. Les régions courtes (~ 5 nucléotides) situées en amont de la boîte C et en aval de la boîte D sont généralement complémentaires des bases et forment une structure tige-boîte, qui rapproche les motifs des boîtes C et D. Il a été démontré que cette structure tige-boîte est essentielle à la synthèse correcte des snoRNA et à la localisation nucléolaire. De nombreux snoRNA à boîte C/D contiennent également une copie supplémentaire moins bien conservée des motifs C et D (appelés C' et D') située dans la partie centrale de la molécule de snoRNA. Une région conservée de 10 à 21 nucléotides en amont de la boîte D est complémentaire du site de méthylation de l'ARN cible et permet au snoRNA de former un duplex d'ARN avec l'ARN. Le nucléotide à modifier dans l'ARN cible est généralement situé à la 5e position en amont de la boîte D (ou de la boîte D'). Les snoRNA à boîte C/D s'associent à quatre protéines évolutives conservées et essentielles - la fibrillarine (Nop1p), NOP56 , NOP58 et SNU13 (protéine de 15,5 kD chez les eucaryotes ; son homologue archéen est L7Ae) - qui constituent le snoRNP à boîte C/D de base.
Il existe un snoRNA eucaryote à boîte C/D ( snoRNA U3 ) dont il n'a pas été démontré qu'il guidait la méthylation 2'- O . Au lieu de cela, il fonctionne dans le traitement de l'ARNr en dirigeant le clivage du pré-ARNr.
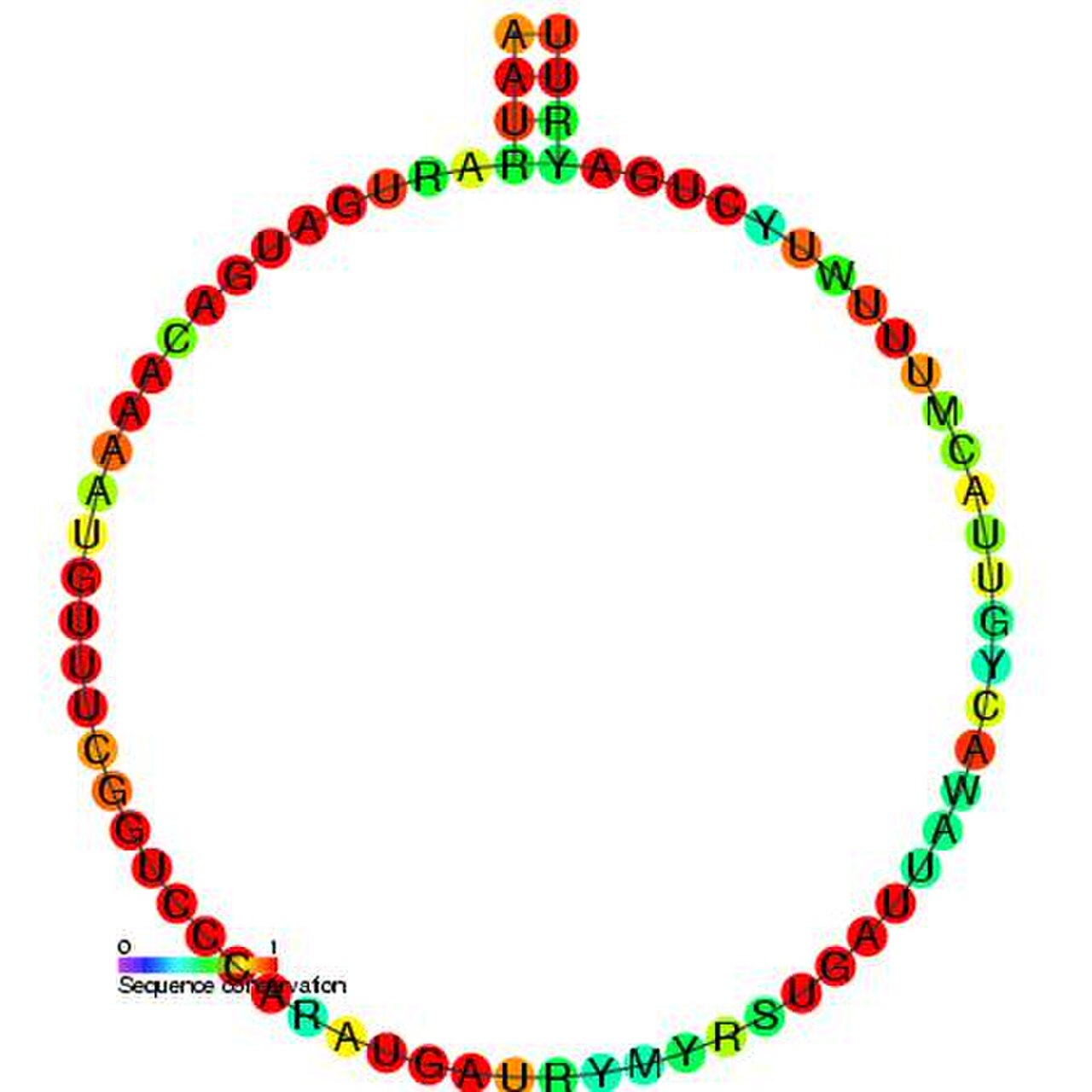
Boîte H/ACA
Les snoRNA à boîte H/ACA ont une structure secondaire commune composée de deux épingles à cheveux et de deux régions monocaténaires appelées structure épingle à cheveux-charnière-épingle à cheveux-queue. Les snoRNA à boîte H/ACA contiennent également des motifs de séquence conservés connus sous le nom de boîte H (consensus ANANNA) et de boîte ACA (ACA). Les deux motifs sont généralement situés dans les régions monocaténaires de la structure secondaire. Le motif H est situé dans la charnière et le motif ACA est situé dans la région de la queue ; à 3 nucléotides de l'extrémité 3' de la séquence. Les régions en épingle à cheveux contiennent des renflements internes appelés boucles de reconnaissance dans lesquelles se trouvent les séquences guides antisens (bases complémentaires de la séquence cible). Ces séquences guides marquent essentiellement l'emplacement de l'uridine sur l'ARNr cible qui va être modifié. Cette séquence de reconnaissance est bipartite (construite à partir des deux bras différents de la région de boucle) et forme des pseudo-nœuds complexes avec l'ARN cible. Français Les snoRNA à boîte H/ACA s'associent à quatre protéines conservées et essentielles au cours de l'évolution - la dyskérine (Cbf5p), GAR1 , NHP2 et NOP10 - qui constituent le cœur du snoRNP à boîte H/ACA. La dyskérine est probablement le composant catalytique du complexe ribonucléoprotéique (RNP) car elle possède plusieurs séquences de pseudouridine synthase conservées et est étroitement liée à la pseudouridine synthase qui modifie l'uridine dans l'ARNt . Dans les cellules eucaryotes inférieures telles que les trypanosomes, des ARN similaires existent sous la forme d'une structure en épingle à cheveux simple et d'une boîte AGA au lieu d'une boîte ACA à l'extrémité 3' de l'ARN. Comme les trypanosomes, Entamoeba histolytica a une population mixte de snoRNA à boîte H/ACA simple et double en épingle à cheveux. Il a été rapporté qu'il y avait eu transformation de l'ARNsno à double épingle à cheveux H/ACA en ARNsno à simple épingle à cheveux. Cependant, contrairement aux trypanosomes, il présente un motif ACA régulier au niveau de la queue 3'. [19]
Le composant ARN de la télomérase humaine (hTERC) contient un domaine H/ACA pour la formation de pré-RNP et la localisation nucléolaire de la RNP de la télomérase elle-même. Le snoRNP H/ACA a été impliqué dans la maladie génétique rare appelée dyskératose congénitale (DKC) en raison de son affiliation avec la télomérase humaine. Les mutations dans le composant protéique du snoRNP H/ACA entraînent une réduction des niveaux physiologiques de TERC. Cela a été fortement corrélé à la pathologie à l'origine de la DKC, qui semble être principalement une maladie due à un mauvais entretien des télomères .
Boîte composite H/ACA et C/D
Un snoRNA U85 guide inhabituel qui fonctionne à la fois dans la méthylation du 2'-O-ribose et la pseudouridylation du petit ARN nucléaire (snRNA) U5 a été identifié. Ce snoRNA composite contient à la fois les domaines de boîte C/D et H/ACA et s'associe aux protéines spécifiques de chaque classe de snoRNA (fibrillarine et Gar1p, respectivement). D'autres snoRNA composites ont maintenant été caractérisés.
Ces snoRNA composites se sont accumulés dans un organite subnucléaire appelé corps de Cajal et sont appelés petits ARN spécifiques du corps de Cajal (scaRNA). Cela contraste avec la majorité des snoRNA à boîte C/D ou à boîte H/ACA, qui se localisent dans le nucléole. Ces ARN spécifiques du corps de Cajal seraient impliqués dans la modification des ARN épissosomiques U1, U2, U4, U5 et U12 transcrits par l'ARN polymérase II. Tous les snoRNA localisés dans les corps de Cajal ne sont pas des snoRNA composites à boîte C/D et H/ACA.
SnoRNA orphelins
Les cibles des snoRNA nouvellement identifiés sont prédites sur la base de la complémentarité de séquence entre les ARN cibles putatifs et les éléments antisens ou les boucles de reconnaissance dans la séquence du snoRNA. Cependant, il existe un nombre croissant de guides « orphelins » sans cible ARN connue, ce qui suggère qu'il pourrait y avoir plus de protéines ou de transcrits impliqués dans l'ARNr qu'auparavant et/ou que certains snoRNA ont des fonctions différentes qui ne concernent pas l'ARNr. Il existe des preuves que certains de ces snoRNA orphelins régulent les transcrits épissés alternativement. SNORD115 à boîte C/D régule l'épissage alternatif de l' ARNm du récepteur de la sérotonine 2C via une région de complémentarité conservée. Un autre snoRNA à boîte C/D, SNORD116 , qui réside dans le même groupe que SNORD115, aurait 23 cibles possibles dans les gènes codant pour des protéines en utilisant une approche bioinformatique . Parmi ceux-ci, une grande fraction s'est avérée être épissée alternativement, suggérant un rôle de SNORD116 dans la régulation de l'épissage alternatif.
Plus récemment, il a été suggéré que SNORD90 pouvait guider les modifications de la N6-méthyladénosine (m6A) sur les transcrits d'ARN cibles. Plus spécifiquement, Lin et al. ont démontré que SNORD90 peut réduire l'expression de la neuréguline 3 (NRG3).
Modifications de la cible
L'effet précis des modifications de méthylation et de pseudouridylation sur la fonction des ARN matures n'est pas encore connu. Les modifications ne semblent pas essentielles mais sont connues pour améliorer subtilement le repliement de l'ARN et l'interaction avec les protéines ribosomales. Pour étayer leur importance, les modifications du site cible sont exclusivement situées dans des domaines conservés et fonctionnellement importants de l'ARN mature et sont généralement conservées parmi les eucaryotes distants. Une nouvelle méthode, Nm-REP-seq, a été développée pour enrichir les 2'-O-méthylations guidées par les snoRNA C/D en utilisant l'exoribonucléase d'ARN (Mycoplasma genitalium RNase R, MgR) et la réactivité d'oxydation du periodate pour éliminer les nucléosides 2'-hydroxylés (2'-OH).
- Le ribose 2′-O-méthylé provoque une augmentation de la conformation 3′-endo
- La pseudouridine (psi/Ψ) ajoute une autre option pour la liaison H.
- L'ARN fortement méthylé est protégé de l'hydrolyse. L'ARNr agit comme un ribozyme en catalysant sa propre hydrolyse et son épissage.
Organisation génomique
Les SnoRNA sont situés de manière diverse dans le génome. La majorité des gènes snoRNA des vertébrés sont codés dans les introns des gènes codant des protéines impliquées dans la synthèse ou la traduction des ribosomes, et sont synthétisés par l'ARN polymérase II . Il a également été démontré que les SnoRNA sont situés dans des régions intergéniques, des ORF de gènes codant des protéines et des UTR. Les SnoRNA peuvent également être transcrits à partir de leurs propres promoteurs par l'ARN polymérase II ou III .
Locaux imprimés
Dans le génome humain, il existe au moins deux exemples où les snoRNA à boîte C/D se trouvent en répétitions en tandem dans des locus imprimés . Ces deux locus (14q32 sur le chromosome 14 et 15q11q13 sur le chromosome 15) ont été largement caractérisés et, dans les deux régions, de multiples snoRNA ont été trouvés situés dans des introns en groupes de copies étroitement liées.
Dans la région 15q11q13, cinq snoRNA différents ont été identifiés ( SNORD64 , SNORD107, SNORD108, SNORD109 (deux copies), SNORD116 (29 copies) et SNORD115 (48 copies). La perte des 29 copies de SNORD116 (HBII-85) de cette région a été identifiée comme une cause du syndrome de Prader-Willi tandis que le gain de copies supplémentaires de SNORD115 a été lié à l'autisme .
La région 14q32 contient des répétitions de deux snoRNA SNORD113 (9 copies) et SNORD114 (31 copies) dans les introns d'un transcrit d'ARNnc spécifique au tissu ( MEG8 ). Il a été démontré que le domaine 14q32 partage des caractéristiques génomiques communes avec les loci 15q11-q13 imprimés et un rôle possible des répétitions en tandem des snoRNA à boîte C/D dans l'évolution ou le mécanisme des loci imprimés a été suggéré.
Autres fonctions
Les snoRNA peuvent fonctionner comme des miRNA . Il a été démontré que l'ACA45 humain est un snoRNA authentique qui peut être transformé en un miRNA mature de 21 nucléotides de long par l' endoribonucléase de la famille RNAse III dicer . Ce produit snoRNA a été précédemment identifié comme mmu-miR-1839 et s'est avéré être traité indépendamment de l'autre endoribonucléase génératrice de miRNA drosha . bioinformatiques ont révélé que des fragments de type miRNA dérivés de snoRNA présumés se produisent dans différents organismes.
Récemment, il a été découvert que les snoRNA peuvent avoir des fonctions non liées à l'ARNr. L'une de ces fonctions est la régulation de l'épissage alternatif du transcrit du gène trans , qui est effectué par le snoRNA HBII-52 , également connu sous le nom de SNORD115.
En novembre 2012, Schubert et al. ont révélé que des ARN spécifiques contrôlent la compaction et l'accessibilité de la chromatine dans les cellules de drosophile .
En juillet 2023, Lin et al. ont montré que les snoRNA ont le potentiel de guider d'autres modifications de l'ARN, en particulier la N6-méthyladénosine , mais cela fait l'objet d'une enquête plus approfondie.
Exemples
TB11Cs4H1
TB11Cs4H1 est un membre de la classe de molécules d'ARN non codant ( ncRNA ) de type H/ACA (un snoRNA) qui guident les sites de modification des uridines en pseudouridines des ARN substrats. Il est prévu que TB11Cs4H1 guide la pseudouridylation de l'ARN ribosomique LSU3 ( rRNA ) au niveau du résidu Ψ1357.