
La zone sous-ventriculaire ( ZSV ) est une région située sur la paroi extérieure de chaque ventricule latéral du cerveau des vertébrés . Elle est présente à la fois dans le cerveau embryonnaire et adulte. Dans la vie embryonnaire, la ZSV fait référence à une zone proliférative secondaire contenant des cellules progénitrices neurales , qui se divisent pour produire des neurones au cours du processus de neurogenèse . Les cellules souches neurales primaires du cerveau et de la moelle épinière, appelées cellules gliales radiales , résident plutôt dans la zone ventriculaire (ZV) (ainsi appelée parce que la ZV tapisse l'intérieur des ventricules en développement ).
Dans le cortex cérébral en développement , qui réside dans le télencéphale dorsal , la SVZ et la VZ sont des tissus transitoires qui n'existent pas chez l'adulte. Cependant, la SVZ du télencéphale ventral persiste tout au long de la vie. La SVZ adulte est composée de quatre couches distinctes d'épaisseur et de densité cellulaire variables ainsi que de composition cellulaire. Avec le gyrus denté de l' hippocampe , la SVZ est l'un des deux endroits où la neurogenèse a été constatée dans le cerveau des mammifères adultes. La neurogenèse de la SVZ adulte prend la forme de précurseurs neuroblastiques d' interneurones qui migrent vers le bulbe olfactif par le flux migratoire rostral . La SVZ semble également être impliquée dans la génération d' astrocytes après une lésion cérébrale.
Structure
Couche I
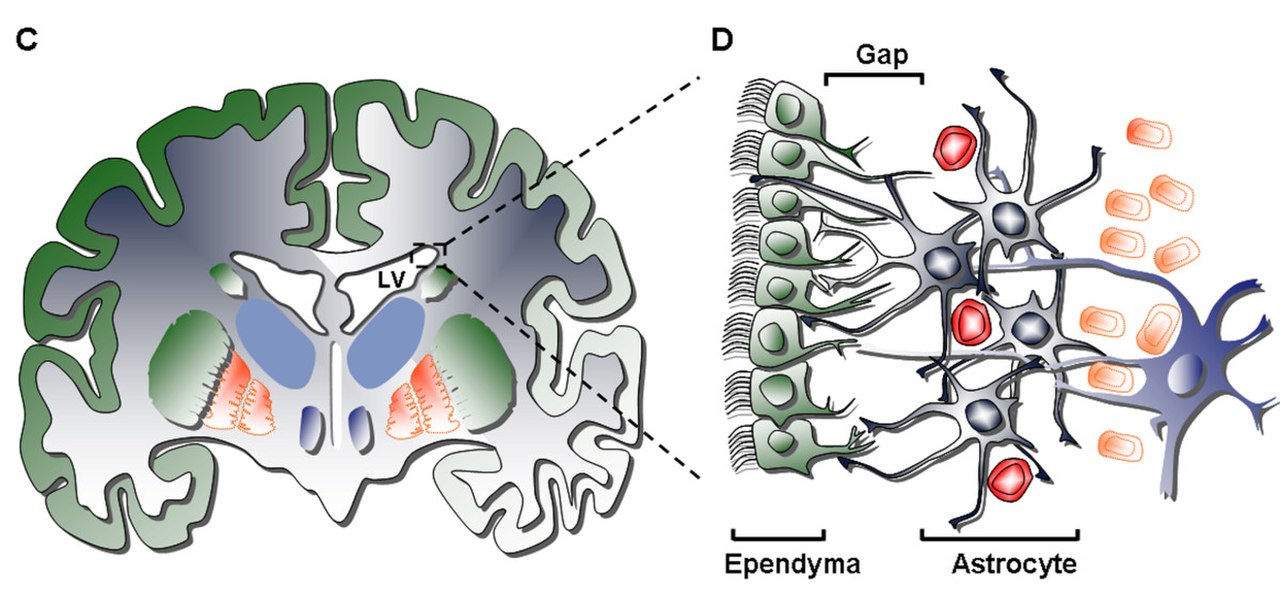
La couche la plus interne (couche I) contient une seule couche (monocouche) de cellules épendymaires tapissant la cavité ventriculaire ; ces cellules possèdent des cils apicaux et plusieurs expansions basales qui peuvent être parallèles ou perpendiculaires à la surface ventriculaire. Ces expansions peuvent interagir intimement avec les processus astrocytaires qui sont interconnectés avec la couche hypocellulaire (couche II).
Couche II
La couche secondaire (couche II) fournit un espace hypocellulaire jouxtant la première et contient un réseau de processus astrocytaires positifs à la protéine acide fibrillaire gliale ( GFAP ) fonctionnellement corrélés qui sont liés à des complexes jonctionnels, mais qui n'ont pas de corps cellulaires à l'exception des rares somas neuronaux. Bien que la fonction de cette couche soit encore inconnue chez l'homme, on a émis l'hypothèse que les interconnexions astrocytaires et épendymaires des couches I et II pourraient agir pour réguler les fonctions neuronales, établir l'homéostasie métabolique et/ou contrôler la prolifération et la différenciation des cellules souches neuronales au cours du développement. Potentiellement, de telles caractéristiques de la couche peuvent agir comme un reste de la vie développementale précoce ou une voie de migration cellulaire étant donné la similitude avec une couche homologue dans la SVZ bovine qui présente des cellules migratoires communes uniquement aux mammifères d'ordre supérieur.
Couche III
La troisième couche (couche III) forme un ruban de corps cellulaires d'astrocytes qui sont censés maintenir une sous-population d'astrocytes capables de proliférer in vivo et de former des neurosphères multipotentes avec des capacités d'auto-renouvellement in vitro. Bien que certains oligodendrocytes et cellules épendymaires aient été trouvés dans le ruban, ils remplissent non seulement une fonction inconnue, mais sont rares par rapport à la population d' astrocytes qui réside dans la couche. Les astrocytes présents dans la couche III peuvent être divisés en trois populations par microscopie électronique , sans fonctions uniques encore reconnaissables ; le premier type est un petit astrocyte de longues projections horizontales et tangentielles principalement trouvées dans la couche II ; le deuxième type se trouve entre les couches II et III ainsi qu'à l'intérieur du ruban d'astrocytes, caractérisé par sa grande taille et ses nombreux organites ; le troisième type se trouve généralement dans les ventricules latéraux juste au-dessus de l' hippocampe et est de taille similaire au deuxième type mais contient peu d'organites.
Couche IV
La quatrième et dernière couche (couche IV) sert de zone de transition entre la couche III avec son ruban d' astrocytes et le parenchyme cérébral . Elle est identifiée par une forte présence de myéline dans la région.
Types de cellules
Quatre types de cellules sont décrits dans le SVZ :
1. Cellules épendymaires ciliées (type E) : elles sont positionnées face à la lumière du ventricule et ont pour fonction de faire circuler le liquide céphalo-rachidien .
2. Neuroblastes prolifératifs (type A) : expriment PSA-NCAM ( NCAM1 ), Tuj1 ( TUBB3 ) et Hu, et migrent en ordre linéaire vers le bulbe olfactif
3. Cellules à prolifération lente (type B) : expriment la nestine et la GFAP et servent à envelopper les neuroblastes de type A en migration
4. Cellules à prolifération active ou progéniteurs amplificateurs de transit (type C) : expriment la nestine et forment des amas espacés parmi les chaînes dans toute la région
Fonction
La SVZ est un site connu de neurogenèse et d'auto-renouvellement des neurones dans le cerveau adulte , en raison des types de cellules en interaction, des molécules extracellulaires et de la régulation épigénétique localisée favorisant cette prolifération cellulaire. Avec la zone sous-granulaire du gyrus denté , la zone sous-ventriculaire sert de source de cellules souches neurales (NSC) dans le processus de neurogenèse adulte . Elle abrite la plus grande population de cellules proliférantes dans le cerveau adulte des rongeurs, des singes et des humains. En 2010, il a été démontré que l'équilibre entre les cellules souches neurales et les cellules progénitrices neurales (NPC) est maintenu par une interaction entre la voie de signalisation du récepteur du facteur de croissance épidermique et la voie de signalisation Notch .
Bien qu'elle n'ait pas encore été étudiée en profondeur dans le cerveau humain, la fonction de la SVZ dans le cerveau des rongeurs a été, dans une certaine mesure, examinée et définie pour ses capacités. Grâce à ces recherches, il a été découvert que l' astrocyte à double fonction est la cellule dominante dans la SVZ des rongeurs ; cet astrocyte agit non seulement comme une cellule souche neuronale, mais aussi comme une cellule de soutien qui favorise la neurogenèse par interaction avec d'autres cellules. Cette fonction est également induite par la microglie et les cellules endothéliales qui interagissent de manière coopérative avec les cellules souches neuronales pour favoriser la neurogenèse in vitro, ainsi que par des composants de la matrice extracellulaire tels que la ténascine-C (aide à définir les limites d'interaction) et Lewis X (lie les facteurs de croissance et de signalisation aux précurseurs neuronaux). La SVZ humaine diffère cependant de la SVZ des rongeurs de deux manières distinctes ; la première est que les astrocytes des humains ne sont pas en étroite juxtaposition avec la couche épendymaire , mais plutôt séparés par une couche dépourvue de corps cellulaires ; la deuxième est que le SVZ humain ne présente pas de chaînes de neuroblastes migrateurs observées dans le SVZ du rongeur, ce qui permet d'obtenir un nombre inférieur de cellules neuronales chez l'humain que chez le rongeur. Pour cette raison, alors que le SVZ du rongeur s'avère être une source précieuse d'informations concernant le SVZ et sa relation structure-fonction, le modèle humain s'avérera sensiblement différent.
Les modifications épigénétiques de l'ADN jouent un rôle central dans la régulation de l'expression des gènes au cours de la différenciation des cellules souches neurales . La conversion de la cytosine en 5-méthylcytosine (5mC) dans l'ADN par l'ADN méthyltransférase DNMT3A semble être un type important de modification épigénétique se produisant dans la SVZ.
De plus, certaines théories actuelles suggèrent que la SVZ pourrait également servir de site de prolifération pour les cellules souches tumorales cérébrales (BTSC), qui sont similaires aux cellules souches neurales dans leur structure et leur capacité à se différencier en neurones , astrocytes et oligodendrocytes . Des études ont confirmé qu'une petite population de BTSC peut non seulement produire des tumeurs, mais aussi les maintenir grâce à un auto-renouvellement inné et à des capacités multipotentes . Bien que cela ne permette pas de déduire que les BTSC proviennent de cellules souches neurales, cela soulève une question intéressante quant à la relation qui existe entre nos propres cellules et celles qui peuvent causer tant de dommages.
Recherches actuelles
De nombreux aspects différents de la SVZ font actuellement l'objet de recherches par des individus des secteurs public et privé. Ces intérêts de recherche vont du rôle de la SVZ dans la neurogenèse , la migration neuronale dirigée, à la tumorigenèse mentionnée précédemment , ainsi que bien d'autres. Vous trouverez ci-dessous des résumés des travaux de trois groupes de laboratoire différents se concentrant principalement sur un aspect de la SVZ ; il s'agit notamment du rôle de la SVZ dans le remplacement cellulaire après une lésion cérébrale, la simulation de la prolifération des cellules souches embryonnaires et son rôle dans divers cancers tumoraux.
Rôle dans le remplacement cellulaire après une lésion cérébrale
Dans leur étude, Romanko et al. ont caractérisé l'impact d'une lésion cérébrale aiguë sur la SVZ. Dans l'ensemble, les auteurs ont déterminé que des lésions modérées de la SVZ permettaient une récupération tandis que des lésions plus graves causaient des dommages permanents à la région. De plus, la population de cellules souches neurales au sein de la SVZ est probablement responsable de cette réponse aux lésions.
Les effets de l'irradiation sur la SVZ ont permis de déterminer la quantité ou la dose de rayonnement pouvant être administrée, en fonction principalement de la tolérance des cellules normales proches de la tumeur . Comme décrit, l'augmentation de la dose de rayonnement et l'âge ont entraîné une diminution de trois types de cellules de la SVZ, mais une capacité de réparation de la SVZ a été observée malgré l'absence de nécrose de la substance blanche ; cela s'est probablement produit parce que la SVZ a pu remplacer progressivement la névroglie du cerveau. Des chimiothérapies ont également été testées pour leurs effets sur la SVZ, car elles sont actuellement utilisées pour de nombreuses maladies, mais entraînent des complications au sein du système nerveux central . Pour ce faire, le méthotrexate (MTX) a été utilisé seul et en combinaison avec la radiothérapie pour constater qu'environ 70 % de la densité nucléaire totale de la SVZ avait été épuisée, mais compte tenu de la perte de cellules neuroblastiques (cellules progénitrices ), il était remarquable de constater que les cellules souches neurales de la SVZ généreraient toujours des neurosphères similaires à celles des sujets qui n'avaient pas reçu un tel traitement. En ce qui concerne l'interruption de l'apport sanguin au cerveau, l'hypoxie / ischémie cérébrale (H/I) a également diminué le nombre de cellules de la SVZ de 20 %, 50 % des neurones du striatum et du néocortex étant détruits, mais les types de cellules de la SVZ tuées étaient aussi hétérogènes que la région elle-même. Lors de tests ultérieurs, il a été constaté qu'une partie différente de chaque cellule était éliminée, mais que la population de cellules de la SVZ médiane restait principalement vivante. Cela peut assurer une certaine résilience de ces cellules, les cellules progénitrices non engagées agissant comme la population proliférante après l'ischémie . Les lésions cérébrales mécaniques induisent également la migration et la prolifération cellulaires, comme cela a été observé chez les rongeurs, et elles peuvent également augmenter le nombre de cellules, ce qui contredit l'idée précédemment admise selon laquelle aucune nouvelle cellule neuronale ne peut être générée.
En conclusion, ce groupe a pu déterminer que les cellules de la SVZ sont capables de produire de nouveaux neurones et de la glie tout au long de la vie, à condition qu'elles ne subissent aucun dommage car elles sont sensibles à tout effet délétère. Par conséquent, la SVZ peut se rétablir après une blessure légère et potentiellement fournir une thérapie cellulaire de remplacement à d'autres régions affectées du cerveau.
Rôle du neuropeptide Y dans la neurogenèse
Dans une tentative de caractériser et d'analyser le mécanisme de prolifération des cellules neuronales dans la zone sous-ventriculaire, Decressac et al. ont observé la prolifération de précurseurs neuronaux dans la zone sous-ventriculaire de la souris par injection du neuropeptide Y (NPY). Le NPY est une protéine couramment exprimée du système nerveux central qui a déjà été montrée comme stimulant la prolifération des cellules neuronales dans l' épithélium olfactif et l'hippocampe . Les effets du peptide ont été observés par marquage BrdU et phénotypage cellulaire qui ont fourni des preuves de la migration des neuroblastes à travers le flux migratoire rostral vers le bulbe olfactif (confirmant les expériences précédentes) et vers le striatum . Ces données soutiennent l'hypothèse de l'auteur selon laquelle la neurogenèse serait stimulée par l'introduction d'un tel peptide .
Le NPY étant un peptide de 36 acides aminés associé à de nombreuses conditions physiologiques et pathologiques, il possède de multiples récepteurs largement exprimés dans le cerveau des rongeurs en développement et matures. Cependant, d'après les études in vivo réalisées par ce groupe, le récepteur Y1 a montré des effets neuroprolifératifs spécifiquement médiés par l'induction du NPY avec une expression accrue dans la zone sous-ventriculaire. L'identification du récepteur Y1 met également en lumière le fait que le phénotype des cellules exprimées à partir de tels événements mitotiques sont en fait des cellules de type DCX + ( neuroblastes qui migrent directement vers le striatum ). Parallèlement aux effets de l'injection de NPY sur les paramètres de la dopamine , du GABA et du glutamate striataux pour réguler la neurogenèse dans la zone sous-ventriculaire (étude précédente), cette découverte est toujours à l'étude car elle pourrait être un modulateur secondaire des neurotransmetteurs susmentionnés .
Comme il est nécessaire pour toute recherche, ce groupe a mené ses expériences avec une perspective large sur l'application de ses résultats, qui, selon eux, pourraient potentiellement bénéficier aux candidats potentiels à la réparation endogène du cerveau par la stimulation de la prolifération des cellules souches neurales de la zone sous-ventriculaire. Cette régulation moléculaire naturelle de la neurogenèse adulte serait complétée par des thérapies à base de molécules appropriées telles que le récepteur NPY et Y1 testé, en plus des dérivés pharmacologiques , pour permettre des formes gérables de troubles neurodégénératifs de la zone striatale.
En tant que source potentielle de tumeurs cérébrales
Dans une tentative de caractériser le rôle de la zone sous-ventriculaire dans la tumorigenèse potentielle , Quinones-Hinojosa et al. ont découvert que les cellules souches tumorales cérébrales (BTSC) sont des cellules souches qui peuvent être isolées des tumeurs cérébrales par des tests similaires à ceux utilisés pour les cellules souches neuronales. En formant des sphères clonales similaires aux neurosphères des cellules souches neuronales, ces BTSC ont pu se différencier en neurones , astrocytes et oligodendrocytes in vitro , mais plus important encore, capables d'initier des tumeurs à de faibles concentrations cellulaires, offrant une capacité d'auto-renouvellement. Il a donc été proposé qu'une petite population de BTSC avec de telles capacités d'auto-renouvellement maintenait des tumeurs dans des maladies telles que la leucémie et le cancer du sein .
Plusieurs facteurs caractéristiques ont conduit à l'idée selon laquelle les cellules souches neuronales (NSC) seraient à l'origine des BTSC, car elles partagent plusieurs caractéristiques. Ces caractéristiques sont présentées dans la figure.
Ce groupe fournit la preuve du rôle apparent de la SVZ dans la tumorigenèse, comme le démontre la possession de récepteurs mitogéniques et leur réponse à la stimulation mitogénique, en particulier les cellules de type C qui expriment le récepteur du facteur de croissance épidermique (EGFR), ce qui les rend hautement prolifératives et invasives. De plus, l'existence de cellules microgliales et endothéliales au sein de la SVZ s'est avérée améliorer la neurogenèse , tout en permettant une migration directionnelle des neuroblastes à partir de la SVZ.
Récemment, la SVZ humaine a été caractérisée chez des patients atteints de tumeurs cérébrales au niveau phénotypique et génétique. Ces données révèlent que chez la moitié des patients, la SVZ est un site exact de tumorigenèse alors que chez les autres patients, elle représente une région infiltrée. Ainsi, il est tout à fait possible que chez l'homme, une relation existe entre la génération de NSC de la région et les cellules auto-renouvelables de tumeurs primaires qui cèdent la place à des tumeurs secondaires une fois retirées ou irradiées.
Bien qu'il reste à prouver avec certitude si les cellules souches SVZ sont les cellules d'origine des tumeurs cérébrales telles que les gliomes, des preuves solides suggèrent une agressivité tumorale et une mortalité accrues chez les patients dont les gliomes de haut grade infiltrent ou entrent en contact avec la SVZ.
Dans le cancer de la prostate, la neurogenèse induite par la tumeur est caractérisée par le recrutement de cellules progénitrices neurales (CPN) à partir de la SVZ. Les CPN infiltrent la tumeur où elles se différencient en neurones autonomes (principalement des neurones adrénergiques) qui stimulent la croissance tumorale.