Démarche
( Apprenez comment et quand supprimer ce message ) Aperçu a été le pionnier de l'analyse scientifique et de la classification des allures. Le mouvement de chaque membre a été dé...
( Apprenez comment et quand supprimer ce message ) Aperçu a été le pionnier de l'analyse scientifique et de la classification des allures. Le mouvement de chaque membre a été dé...
cycle complet dans un intervalle de temps identique ; autrement, la relation entre les membres peut varier et un schéma stable ne peut se dégager. Ainsi, toute allure peut être entièrement décrite en fonction du début et de la fin de la phase d'appui de trois membres par rapport au cycle d'un membre de référence, généralement le membre postérieur gauche .
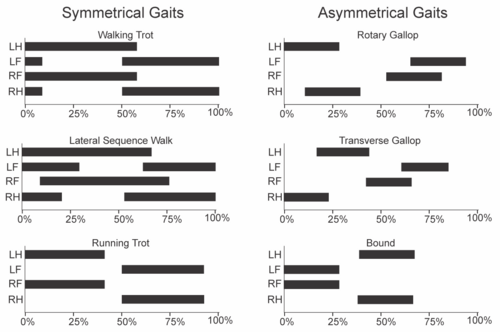
Les allures sont généralement classées en « symétriques » et « asymétriques » selon le mouvement des membres. Ces termes n'ont rien à voir avec la symétrie gauche-droite . Dans une allure symétrique, les membres gauche et droit d'une paire alternent, tandis que dans une allure asymétrique, les membres se meuvent simultanément. Les allures asymétriques sont parfois qualifiées d'« allures bondissantes », en raison de la présence d'une phase d'arrêt.
Les variables clés de la démarche sont le facteur d'appui et le rapport de phase entre les membres antérieurs et postérieurs. Le facteur d'appui correspond au pourcentage du cycle total pendant lequel un membre donné est en contact avec le sol. Cette valeur est généralement identique pour les membres antérieurs et postérieurs, sauf si l'animal se déplace selon une démarche spécialement entraînée ou s'il accélère ou décélère . Un facteur d'appui supérieur à 50 % correspond à la marche, tandis qu'un facteur inférieur à 50 % correspond à la course. Le rapport de phase entre les membres antérieurs et postérieurs représente la relation temporelle entre ces deux membres. Si les membres antérieurs et postérieurs du même côté entament la phase d'appui simultanément, le rapport de phase est de 0 (ou 100 %). Si le membre antérieur du même côté touche le sol une demi-période plus tard que le membre postérieur, le rapport de phase est de 50 %.
Le choix de la locomotion peut avoir des effets qui dépassent les simples modifications de la vitesse et des mouvements des membres, notamment en termes de ventilation . Dépourvus de diaphragme , les lézards et les salamandres doivent dilater et contracter leur paroi corporelle pour faire entrer et sortir l'air de leurs poumons. Or, ce sont les mêmes muscles qui servent à onduler latéralement le corps lors de la locomotion. Ils ne peuvent donc pas se déplacer et respirer simultanément, une situation appelée contrainte de Carrier . Certains, comme les varans , peuvent toutefois contourner cette restriction grâce à la respiration buccale . À l'inverse, la flexion de la colonne vertébrale d'un mammifère au galop permet aux viscères abdominaux d'agir comme un piston, gonflant et dégonflant les poumons au rythme des mouvements de la colonne vertébrale, ce qui améliore la ventilation et favorise les échanges gazeux .
Les animaux utilisent généralement différentes allures en fonction de leur vitesse. Presque tous les animaux sont capables d'allures symétriques, tandis que les allures asymétriques sont principalement l'apanage des mammifères, qui peuvent effectuer une flexion vertébrale suffisante pour augmenter la longueur de leur foulée (bien que les petits crocodiliens soient capables d'une allure bondissante). Les allures à séquence latérale lors de la marche et de la course sont les plus fréquentes chez les mammifères[3], mais les mammifères arboricoles tels que les singes, certains opossums et les kinkajous utilisent des allures à séquence diagonale pour une meilleure stabilité[3]. Les marches et courses à séquence diagonale (aussi appelées trot) sont le plus souvent utilisées par les tétrapodes rampants tels que les salamandres et les lézards, en raison des oscillations latérales de leur corps lors du mouvement. Les bipèdes constituent un cas particulier, et la plupart d'entre eux n'utilisent que trois allures — la marche, la course et le saut — lors de leurs déplacements naturels. Les autres allures, comme le saut à cloche-pied chez l'humain, ne sont pas utilisées sans effort volontaire.
Les locomotions des hexapodes ont été bien caractérisées, notamment chez la drosophile et les phasmes (Phasmatodea). La drosophile utilise une locomotion tripode où trois pattes se balancent simultanément tandis que les trois autres restent au sol en appui. Cependant, la variabilité de la locomotion est continue. Les mouches ne présentent pas de transitions distinctes entre les locomotions, mais sont plus susceptibles de marcher en configuration tripode à des vitesses élevées. À des vitesses plus faibles, elles sont plus susceptibles de marcher avec quatre ou cinq pattes en appui. La coordination tétrapode (lorsque les quatre pattes sont en appui) se caractérise par le balancement simultané de paires de pattes diagonalement opposées. La marche ondulatoire (parfois appelée marche métachrone) décrit une marche où une seule patte entre en phase de balancement à la fois. Ce mouvement se propage de l'arrière vers l'avant d'un côté du corps, puis de l'autre. Le phasme, un hexapode de plus grande taille, n'adopte une locomotion tripode que durant le stade larvaire. À l'âge adulte et à faible vitesse, ils marchent généralement selon une onde métachrone, où une seule patte se balance à la fois. À vitesse plus élevée, ils marchent selon une coordination tétrapode, avec deux pattes appariées en mouvement oscillant ou selon une onde métachrone, ne déplaçant qu'une seule patte à la fois.
Si les allures peuvent être classées selon le type de foulée, de nouvelles études intégrant la cinématique corporelle et l'enregistrement des forces ont permis d'établir une classification alternative, basée sur la mécanique du mouvement . Dans ce système, les mouvements sont divisés en marche et course. La marche est caractérisée par un mouvement de propulsion du corps au-dessus des jambes, souvent décrit comme un pendule inversé (présentant des fluctuations d' énergie cinétique et potentielle déphasées), un mécanisme décrit par Giovanni Cavagna . Lors de la course, l'énergie cinétique et potentielle fluctuent en phase, et la variation d'énergie est transmise aux muscles , aux os , aux tendons et aux ligaments qui agissent comme des ressorts (elle est ainsi décrite par le modèle masse-ressort ).

La vitesse détermine généralement le choix de l'allure : les mammifères quadrupèdes passent de la marche à la course, puis au galop, à mesure que leur vitesse augmente. Chaque allure possède une vitesse optimale, à laquelle la consommation de calories par mètre est minimale ; le coût énergétique augmente à des vitesses inférieures ou supérieures. Les transitions d'allure se produisent à une vitesse proche de celle où le coût d'une marche rapide devient supérieur à celui d'une course lente. Les animaux en liberté se déplacent généralement à la vitesse optimale pour leur allure afin de minimiser leur dépense énergétique. Le coût du transport est utilisé pour comparer l'énergétique des différentes allures, ainsi que celle des allures de différentes espèces animales.
Malgré les différences de nombre de pattes observées chez les vertébrés terrestres , selon le modèle du pendule inversé pour la marche et le modèle masse-ressort pour la course, on observe des allures de marche et de course chez les animaux à 2, 4, 6 pattes ou plus. Le terme « allure » a même été appliqué aux organismes volants et nageurs qui produisent des sillages tourbillonnaires distincts .