Les lymphocytes T à mémoire sont un sous-ensemble de lymphocytes T qui pourraient avoir certaines des mêmes fonctions que les lymphocytes B à mémoire . Leur lignée n'est pas claire.
Fonction
Les lymphocytes T à mémoire spécifiques d'antigènes spécifiques aux virus ou à d'autres molécules microbiennes peuvent être trouvés dans les sous-ensembles de lymphocytes T à mémoire centrale (T CM ) et de lymphocytes T à mémoire effectrice (T EM ) . Bien que la plupart des informations soient actuellement basées sur des observations dans le sous-ensemble de lymphocytes T cytotoxiques ( CD8 -positifs), des populations similaires semblent exister pour les lymphocytes T auxiliaires ( CD4 -positifs) et les lymphocytes T cytotoxiques. La fonction principale des cellules à mémoire est d'augmenter la réponse immunitaire après la réactivation de ces cellules par la réintroduction du pathogène concerné dans l'organisme. Il est important de noter que ce domaine fait l'objet d'études intensives et que certaines informations peuvent ne pas être disponibles à ce jour.
- Cellules T à mémoire centrale (T CM ) : les lymphocytes T CM ont plusieurs attributs en commun avec les cellules souches, le plus important étant la capacité d'auto-renouvellement, principalement en raison du niveau élevé de phosphorylation du facteur de transcription clé STAT5 . Chez la souris, les T CM se sont avérés conférer une immunité plus puissante contre les virus, les bactéries et les cellules cancéreuses, par rapport aux lymphocytes T EM dans plusieurs modèles expérimentaux.
- Cellules T effectrices à mémoire (T EM ) : les lymphocytes T EM et T EMRA sont principalement actifs en tant que variantes CD8, étant ainsi principalement responsables de l'action cytotoxique contre les agents pathogènes.
- Cellule T mémoire résidente dans les tissus (T RM ) : Étant donné que les lymphocytes T RM sont présents pendant de longues périodes dans les tissus, ou plus important encore, dans les tissus barrières (l'épithélium par exemple), ils sont essentiels pour une réponse rapide à la rupture de la barrière et à tout pathogène pertinent présent. L'un des mécanismes utilisés par T RM pour restreindre les pathogènes est la sécrétion de granzyme B.
- Cellules T à mémoire de type cellules souches (T SCM ) : Ces lymphocytes sont capables de s'auto-renouveler comme les lymphocytes T CM et sont également capables de générer les sous-populations T CM et T EM . La présence de cette population chez l'homme est actuellement à l'étude.
- Cellule T à mémoire virtuelle (T VM ) : à l'heure actuelle, la seule fonction apparente des cellules T VM est la production de diverses cytokines, mais il existe des spéculations sur leur influence dans la maîtrise des états immunologiques indésirables et leur utilisation dans le traitement des troubles auto-immuns.
Maintien homéostatique
Les clones de lymphocytes T mémoire exprimant un récepteur spécifique des lymphocytes T peuvent persister pendant des décennies dans notre corps. Étant donné que les lymphocytes T mémoire ont une demi-vie plus courte que les lymphocytes T naïfs, la réplication et le remplacement continus des anciennes cellules sont probablement impliqués dans le processus de maintenance. Actuellement, le mécanisme de maintenance des lymphocytes T mémoire n'est pas entièrement compris. L'activation par le récepteur des lymphocytes T peut jouer un rôle. On a découvert que les lymphocytes T mémoire peuvent parfois réagir à de nouveaux antigènes, potentiellement en raison de la diversité et de l'étendue intrinsèques des cibles de liaison des récepteurs des lymphocytes T. Ces lymphocytes T pourraient réagir de manière croisée avec des antigènes environnementaux ou résidents dans notre corps (comme les bactéries dans notre intestin) et proliférer. Ces événements aideraient à maintenir la population de lymphocytes T mémoire. Le mécanisme de réactivité croisée peut être important pour les lymphocytes T mémoire dans les tissus muqueux puisque ces sites ont une densité antigénique plus élevée. Pour ceux qui résident dans le sang, la moelle osseuse, les tissus lymphoïdes et la rate, les cytokines homéostatiques (y compris l'IL-17 et l'IL-15 ) ou la signalisation du complexe majeur d'histocompatibilité II (MHCII) peuvent être plus importantes.
Aperçu de la vie
Les lymphocytes T à mémoire subissent différents changements et jouent différents rôles à différentes étapes de la vie des humains. À la naissance et dans la petite enfance, les lymphocytes T du sang périphérique sont principalement des lymphocytes T naïfs. Grâce à une exposition fréquente aux antigènes, la population de lymphocytes T à mémoire s'accumule. Il s'agit de la phase de génération de la mémoire, qui dure de la naissance à environ 20-25 ans, lorsque notre système immunitaire rencontre le plus grand nombre de nouveaux antigènes. Au cours de la phase d'homéostasie de la mémoire qui vient ensuite, le nombre de lymphocytes T à mémoire atteint un plateau et est stabilisé par le maintien homéostatique. À ce stade, la réponse immunitaire se déplace davantage vers le maintien de l'homéostasie puisque peu de nouveaux antigènes sont rencontrés. La surveillance des tumeurs devient également importante à ce stade. À des stades ultérieurs de la vie, vers 65-70 ans, arrive la phase d' immunosénescence , au cours de laquelle on observe une dysrégulation immunitaire, un déclin de la fonction des lymphocytes T et une sensibilité accrue aux agents pathogènes.
Débat sur la lignée
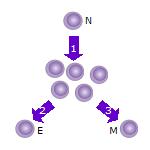
- Après que le lymphocyte T naïf (N) rencontre un antigène, il devient activé et commence à proliférer ( se diviser ) en de nombreux clones ou cellules filles.
- Certains clones de cellules T se différencieront en cellules T effectrices (E) qui exécuteront la fonction de cette cellule (par exemple, produire des cytokines dans le cas des cellules T auxiliaires ou provoquer la destruction cellulaire dans le cas des cellules T cytotoxiques ).
- Certaines cellules formeront des cellules T mémoires (M) qui survivront dans un état inactif dans l’hôte pendant une longue période jusqu’à ce qu’elles rencontrent à nouveau le même antigène et se réactivent.
En avril 2020, la relation de lignée entre les cellules T effectrices et les cellules T mémoires n'était pas claire. Deux modèles concurrents existent. L'un est appelé le modèle On-Off-On. Lorsque les cellules T naïves sont activées par la liaison du récepteur des cellules T (TCR) à l'antigène et à sa voie de signalisation en aval, elles prolifèrent activement et forment un grand clone de cellules effectrices. Les cellules effectrices subissent une sécrétion active de cytokines et d'autres activités effectrices. Après l'élimination de l'antigène , certaines de ces cellules effectrices forment des cellules T mémoires, soit de manière aléatoire, soit sélectionnées en fonction de leur spécificité supérieure. Ces cellules passeraient du rôle d'effecteur actif à un état plus similaire aux cellules T naïves et seraient à nouveau « activées » lors de la prochaine exposition à l'antigène. Ce modèle prédit que les cellules T effectrices peuvent se transformer en cellules T mémoires et survivre, en conservant la capacité de proliférer. Il prédit également que certains profils d'expression génétique suivraient le modèle on-off-on pendant les stades naïf, effecteur et mémoire. Les preuves à l'appui de ce modèle incluent la découverte de gènes liés à la survie et au retour à la maison qui suivent le modèle d'expression on-off-on, notamment le récepteur de l'interleukine-7 alpha (IL-7Rα), Bcl-2, CD26L et d'autres.

dans ce modèle, les cellules T mémoires génèrent des cellules T effectrices, et non l’inverse.
L'autre modèle est le modèle de différenciation développementale. Ce modèle soutient que les cellules effectrices produites par les cellules T naïves hautement activées subiraient toutes une apoptose après l'élimination de l'antigène. Les cellules T mémoires sont plutôt produites par des cellules T naïves qui sont activées mais ne sont jamais entrées avec toute leur force dans le stade effecteur. La progéniture des cellules T mémoires n'est pas complètement activée car elle n'est pas aussi spécifique à l'antigène que les cellules T effectrices en expansion. Des études examinant l'historique de la division cellulaire ont révélé que la longueur du télomère et l'activité de la télomérase étaient réduites dans les cellules T effectrices par rapport aux cellules T mémoires, ce qui suggère que les cellules T mémoires n'ont pas subi autant de division cellulaire que les cellules T effectrices, ce qui est incompatible avec le modèle On-Off-On. Une stimulation antigénique répétée ou chronique des cellules T, comme l'infection par le VIH , induirait des fonctions effectrices élevées mais réduirait la mémoire. Il a également été constaté que les cellules T à prolifération massive sont plus susceptibles de générer des cellules effectrices à courte durée de vie, tandis que les cellules T à prolifération minimale formeraient des cellules à durée de vie plus longue.
Modifications épigénétiques
Les modifications épigénétiques sont impliquées dans le changement des cellules T naïves. Par exemple, dans les cellules T à mémoire CD4 + , les modifications positives des histones marquent les gènes clés des cytokines qui sont régulés à la hausse pendant la réponse immunitaire secondaire, notamment l'IFNγ , l'IL4 et l'IL17A . Certaines de ces modifications ont persisté après l'élimination de l'antigène, établissant une mémoire épigénétique qui permet une activation plus rapide lors de la rencontre avec l'antigène. Pour les cellules T à mémoire CD8 + , certains gènes effecteurs, tels que l'IFNγ , ne seraient pas exprimés mais ils sont prêts transcriptionnellement à s'exprimer rapidement lors de l'activation. De plus, l'amélioration de l'expression de certains gènes dépend également de la force de la signalisation TCR initiale pour la progéniture des cellules T à mémoire, qui est corrélée à l'activation de l'élément régulateur qui modifie directement le niveau d'expression des gènes.
Sous-populations
Historiquement, on pensait que les lymphocytes T à mémoire appartenaient soit aux sous-types effecteurs (cellules T EM ) soit aux sous-types de mémoire centrale (cellules T CM ), chacun ayant son propre ensemble distinctif de marqueurs de surface cellulaire (voir ci-dessous). Par la suite, de nombreuses populations supplémentaires de lymphocytes T à mémoire ont été découvertes, notamment les lymphocytes T à mémoire résidente dans les tissus (T RM ), les lymphocytes T à mémoire souche SCM et les lymphocytes T à mémoire virtuelle . Le thème unificateur unique de tous les sous-types de lymphocytes T à mémoire est qu'ils ont une longue durée de vie et peuvent rapidement se multiplier en un grand nombre de lymphocytes T effecteurs lors d'une réexposition à leur antigène apparenté. Par ce mécanisme, ils fournissent au système immunitaire une « mémoire » contre les agents pathogènes rencontrés précédemment. Les lymphocytes T à mémoire peuvent être CD4 + ou CD8 + et expriment généralement CD45RO tout en étant dépourvus de CD45RA.
Sous-types de cellules T à mémoire
- Les lymphocytes T à mémoire centrale (cellules T CM ) expriment CD45RO, le récepteur de chimiokine CC de type 7 (CCR7) et la L-sélectine (CD62L). Les lymphocytes T à mémoire centrale ont également une expression intermédiaire à élevée de CD44 . Cette sous-population de mémoire se trouve généralement dans les ganglions lymphatiques et dans la circulation périphérique.
- Les cellules T effectrices à mémoire (cellules T EM ) expriment CD45RO mais n'expriment pas CCR7 et L-sélectine . Elles ont également une expression intermédiaire à élevée de CD44 . Comme ces cellules T à mémoire ne possèdent pas de récepteurs CCR7 de localisation des ganglions lymphatiques, on les trouve dans la circulation périphérique et dans les tissus. T EMRA signifie cellules effectrices à mémoire terminale différenciées réexprimant CD45RA, un marqueur généralement présent sur les cellules T naïves.
- Le sous-type de cellules T à mémoire périphérique (cellules T PM ) a été identifié sur la base de l'expression intermédiaire de CX3CR1. Ces cellules peuvent migrer vers les tissus à partir du sang et circuler vers les ganglions lymphatiques d'une manière indépendante de CD62L, afin de surveiller les tissus.
- Les cellules T mémoires résidentes dans les tissus (T RM ) occupent les tissus (peau, poumon, tube digestif, etc.) sans recirculer. Certains marqueurs de surface cellulaire qui ont été associés à T RM sont CD69 et l'intégrine αeβ7 (CD103). Cependant, il convient de noter que les cellules T RM trouvées dans différents tissus expriment différents ensembles de marqueurs de surface cellulaire. RM CD103+ sont localisées de manière restreinte dans les tissus épithéliaux et neuronaux, les cellules T RM localisées dans les glandes salivaires, le pancréas et les voies reproductrices femelles chez la souris n'expriment ni CD69 ni CD103. On pense que les cellules T RM jouent un rôle majeur dans l'immunité protectrice contre les agents pathogènes. Des études ont également suggéré un double rôle des cellules T RM dans la protection et la régulation. Par rapport aux cellules T EM , les cellules T RM sécrètent des niveaux plus élevés de cytokines liées à l'immunité protectrice et expriment des niveaux plus faibles du marqueur de prolifération Ki67. Il a été suggéré que ces caractéristiques pourraient aider au maintien à long terme des cellules T RM , ainsi qu'à maintenir un équilibre entre une réponse rapide à l'invasion antigénique et l'évitement de dommages tissulaires inutiles. RM dysfonctionnelles ont été impliquées dans des maladies auto-immunes, telles que le psoriasis , la polyarthrite rhumatoïde et les maladies inflammatoires chroniques de l'intestin . RM sont spécifiques aux gènes impliqués dans le métabolisme lipidique , étant très actifs, environ 20 à 30 fois plus actifs que dans les autres types de cellules T.
- Les cellules T à mémoire virtuelle (TVM ) diffèrent des autres sous-ensembles de mémoire en ce qu'elles ne naissent pas à la suite d'un événement d'expansion clonale important. Ainsi, bien que cette population dans son ensemble soit abondante dans la circulation périphérique, les clones individuels de cellules T à mémoire virtuelle résident à des fréquences relativement faibles. Une théorie est que la prolifération homéostatique donne naissance à cette population de cellules T. Bien que les cellules T à mémoire virtuelle CD8 aient été les premières à être décrites, on sait maintenant que les cellules à mémoire virtuelle CD4 existent également.
De nombreuses autres sous-populations de lymphocytes T à mémoire ont été suggérées. Les chercheurs ont étudié les cellules souches T à mémoire SCM. Comme les lymphocytes T naïfs, les cellules T SCM sont CD45RO− , CCR7 + , CD45RA+, CD62L+ ( L-sélectine ), CD27+, CD28+ et IL-7Rα+, mais elles expriment également de grandes quantités de CD95, IL-2Rβ, CXCR3 et LFA-1, et présentent de nombreux attributs fonctionnels distinctifs des cellules à mémoire.
Activation indépendante du TCR (témoin)
Les lymphocytes T possèdent la capacité d'être activés indépendamment de leur stimulation par un antigène apparenté, c'est-à-dire sans stimulation du TCR. Aux premiers stades de l'infection, les lymphocytes T spécifiques d'un antigène non apparenté ne sont activés que par la présence d'une inflammation. Cela se produit dans le milieu inflammatoire résultant d'une infection microbienne, d'un cancer ou d'une auto-immunité chez la souris et l'homme et se produit localement ainsi que systématiquement . De plus, les lymphocytes T activés par un témoin peuvent migrer vers le site de l'infection, en raison de l'augmentation de l'expression de CCR5 .
Ce phénomène a été observé principalement dans les cellules T CD8+ à mémoire, qui ont une sensibilité plus faible à la stimulation par les cytokines que leurs homologues naïves et s'activent de cette manière plus facilement. Les cellules T CD8+ à mémoire virtuelle présentent également une sensibilité accrue à l'activation induite par les cytokines dans les modèles murins, mais cela n'a pas été directement démontré chez l'homme. À l'inverse, l'activation indépendante du TCR des cellules T CD8+ naïves reste controversée.
Outre les infections, l'activation par les témoins joue également un rôle important dans l'immunité antitumorale. Dans les tissus cancéreux humains, un nombre élevé de cellules T CD8+ spécifiques du virus, et non spécifiques de la tumeur, a été détecté. Ce type d'activation est considéré comme bénéfique pour l'hôte en termes d'efficacité d'élimination du cancer.
Facteurs d'activation des témoins
Français Les principaux facteurs d'activation des témoins sont les cytokines , telles que l'IL-15 , l'IL-18 , l'IL-12 ou les IFN de type I, qui fonctionnent souvent en synergie. L'IL-15 est responsable de l'activité cytotoxique des cellules T activées par les témoins. Elle induit l' expression de NKG2D (un récepteur généralement exprimé sur les cellules NK ) sur les cellules T CD8+ mémoire, conduisant à une cytotoxicité de type innée, c'est-à-dire la reconnaissance des ligands NKG2D comme indicateurs d'infection, de stress cellulaire et de transformation cellulaire ainsi que de destruction de cellules altérées de manière similaire à celle des cellules NK. Il a été démontré que l'activation du TCR abroge l'expression de NKG2D médiée par l'IL-15 sur les cellules T. De plus, l'IL-15 induit l'expression de molécules cytolytiques, l'expansion cellulaire et améliore la réponse cellulaire à l'IL-18. L'IL-18 est une autre cytokine impliquée dans ce processus, agissant généralement en synergie avec l'IL-12, améliorant la différenciation des cellules T mémoires en cellules effectrices, c'est-à-dire qu'elle induit la production d'IFN-γ et la prolifération cellulaire. Les récepteurs de type Toll (TLR), en particulier le TLR2 , ont également été liés à l'activation indépendante du TCR des cellules T CD8+ lors d'une infection bactérienne.
Activation par un témoin des cellules T CD4+
Français Bien que l'activation indépendante du TCR soit étudiée plus en profondeur dans les cellules T CD8+, il existe une preuve claire de la survenue de ce phénomène dans les cellules T CD4+ . Cependant, il est considéré comme moins efficace, probablement en raison d'une expression plus faible de CD122 (également connu sous le nom d'IL2RB ou IL15RB). De la même manière que leurs homologues CD8+, les cellules T CD4+ mémoire et effectrices présentent une sensibilité accrue à l'activation indépendante du TCR. L'IL-1β , en synergie avec l'IL-12 et l'IL-23, stimule les cellules T CD4+ mémoire et entraîne la réponse Th17 . De plus, l'IL-18, l'IL-12 et l'IL-27 induisent l'expression de cytokines dans les cellules T CD4+ effectrices et mémoires et l'IL-2 est considérée comme un puissant inducteur d'activation des cellules T CD4+ qui peut remplacer la stimulation du TCR même dans les cellules naïves. Il a également été signalé que TLR2 était présent sur les cellules T CD4+ mémoire, qui répondent à leur agoniste par la production d'IFNγ, même sans stimulation du TCR.
Rôle dans la pathogénicité
Bystander activation plays role in the elimination of the spread of infection in its early stages and helps in tumor clearance. However, this type of activation can also have deleterious outcome, especially in chronic infections and autoimmune diseases. Liver injury during chronic Hepatitis B virus infection is a result of non-HBV-specific CD8+ T cell infiltration into the tissue. A similar situation occurs during the acute Hepatitis A virus infection and activated virus unrelated CD4+ T cells contribute to ocular lesions in Herpes Simplex Virus infections.
Increased IL-15 expression and subsequent excessive NKG2D expression was linked to exacerbation of some autoimmune disorders, such as, type I diabetes, multiple sclerosis and inflammatory bowel diseases, for instance Crohn's disease and celiac disease. Furthermore, enhanced TLR2 expression was observed in joints, cartilage and bones of rheumatoid arthritis patients and the presence of its ligand, peptidoglycan, was detected in their synovial fluid.