métabolisme ( / , du , « changement » désigne l’ensemble des chimiques à vie qui se produisent au sein des organismes vivants . trois fonctions principales sont la conversion de l’ énergie une forme utilisable par les cellules ; la transformation des aliments en éléments constitutifs des macromolécules ( biopolymères ) tels que les protéines , les lipides , les acides nucléiques et certains glucides ; et l’élimination des déchets métaboliques . Ces réactions enzymatiques permettent aux organismes de croître, de se reproduire, de maintenir leurs structures et de réagir à leur environnement. Le terme « métabolisme » peut également désigner toutes les réactions chimiques qui se produisent dans les organismes vivants, y compris la digestion et le transport des substances à l’intérieur et entre les différentes cellules. Au sens large, l’ensemble des réactions se produisant à l’intérieur des cellules est appelé métabolisme intermédiaire.
Les réactions métaboliques peuvent être classées en deux catégories : les réactions cataboliques ( dégradation des composés, par exemple la transformation du glucose en pyruvate par la respiration cellulaire ) et les réactions anaboliques (synthèse des composés, tels que les protéines, les glucides, les lipides et les acides nucléiques). Généralement, le catabolisme libère de l’énergie, tandis que l’anabolisme en consomme.
Les réactions chimiques du métabolisme sont organisées en voies métaboliques , au cours desquelles une molécule est transformée en une autre par une série d'étapes, chaque étape étant catalysée par une enzyme spécifique . Les enzymes sont essentielles au métabolisme car elles permettent aux organismes de réaliser des réactions nécessitant de l'énergie et qui ne se produiraient pas spontanément, en les couplant à des réactions spontanées libérant de l'énergie. Les enzymes agissent comme catalyseurs – elles accélèrent une réaction – et permettent également de réguler la vitesse d'une réaction métabolique, par exemple en réponse à des changements dans l' environnement cellulaire ou à des signaux provenant d'autres cellules.
Le système métabolique d'un organisme donné détermine quelles substances lui sont nutritives et lesquelles lui sont toxiques . Par exemple, certains procaryotes utilisent le sulfure d'hydrogène (H₂S) comme nutriment, alors que ce gaz est toxique pour certains animaux. Le sulfure d'hydrogène étant un gazotransmetteur , certains mammifères, dont l'être humain, en produisent naturellement en très faibles concentrations, où il joue un rôle essentiel dans la signalisation et la régulation. Le métabolisme basal d'un organisme correspond à la quantité d'énergie consommée par l'ensemble de ces réactions chimiques.
Une caractéristique remarquable du métabolisme est la similarité des voies métaboliques fondamentales chez des espèces très différentes. Par exemple, l'ensemble des acides carboxyliques , mieux connus comme intermédiaires du cycle de l'acide citrique, est présent chez tous les organismes connus, qu'il s'agisse de la bactérie unicellulaire Escherichia coli ( E. coli) ou d'organismes multicellulaires de grande taille comme les éléphants . Ces similarités dans les voies métaboliques sont probablement dues à leur apparition précoce dans l'histoire de l'évolution , et leur maintien est probablement dû à leur efficacité . Dans diverses maladies, telles que le diabète de type II , le syndrome métabolique et le cancer , le métabolisme normal est perturbé. Le métabolisme des cellules cancéreuses diffère également de celui des cellules normales, et ces différences peuvent être exploitées pour identifier des cibles thérapeutiques dans le traitement du cancer.
Principaux composés biochimiques
Acides aminés et protéines
Lipides
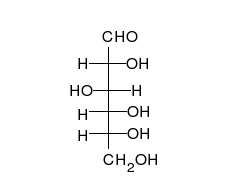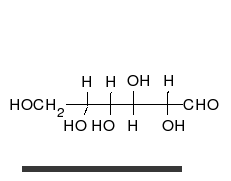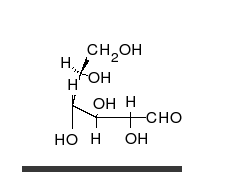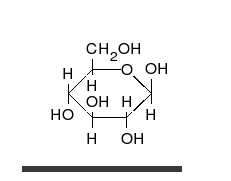
Glucides
Nucléotides
coenzymes
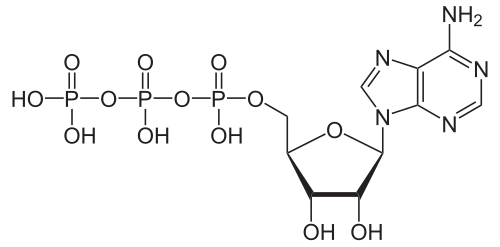
L' adénosine triphosphate (ATP) , coenzyme essentielle, est la monnaie énergétique des cellules. Ce nucléotide sert au transfert d'énergie chimique entre différentes réactions. Les cellules ne contiennent qu'une faible quantité d'ATP, mais comme elle est constamment régénérée, le corps humain peut utiliser chaque jour une quantité d'ATP équivalente à son propre poids. L'ATP fait le lien entre le catabolisme et l'anabolisme . Le catabolisme dégrade les molécules, tandis que l'anabolisme les synthétise. Les réactions cataboliques produisent de l'ATP, et les réactions anaboliques le consomment. L'ATP sert également de transporteur de groupements phosphate lors des réactions de phosphorylation .
Une vitamine est un composé organique nécessaire en petites quantités et que les cellules ne peuvent synthétiser. En nutrition humaine , la plupart des vitamines fonctionnent comme coenzymes après modification ; par exemple, toutes les vitamines hydrosolubles sont phosphorylées ou liées à des nucléotides lorsqu’elles sont utilisées dans les cellules. Le nicotinamide adénine dinucléotide (NAD + ), un dérivé de la vitamine B3 ( niacine ) , est une coenzyme importante qui agit comme accepteur d’hydrogène. Des centaines de déshydrogénases différentes extraient des électrons de leurs substrats et réduisent le NAD + en NADH. Cette forme réduite de la coenzyme est ensuite un substrat pour les réductases cellulaires qui doivent transférer des atomes d’hydrogène à leurs substrats. Le nicotinamide adénine dinucléotide existe sous deux formes apparentées dans la cellule : NADH et NADPH. La forme NAD + /NADH est prédominante dans les réactions cataboliques, tandis que la forme NADP + /NADPH est utilisée dans les réactions anaboliques.
Minéraux et cofacteurs
Les éléments inorganiques abondants agissent comme électrolytes . Les ions les plus importants sont le sodium , le potassium , le calcium , le magnésium , le chlorure , le phosphate et l'ion organique bicarbonate . Le maintien de gradients ioniques précis à travers les membranes cellulaires assure la stabilité de la pression osmotique et du pH . Les ions sont également essentiels au fonctionnement des nerfs et des muscles , car les potentiels d'action dans ces tissus sont générés par l'échange d'électrolytes entre le liquide extracellulaire et le cytosol , le liquide intracellulaire . Les électrolytes entrent et sortent des cellules grâce à des protéines membranaires appelées canaux ioniques . Par exemple, la contraction musculaire dépend du passage du calcium, du sodium et du potassium à travers les canaux ioniques de la membrane cellulaire et des tubules T.
Les métaux de transition sont généralement présents à l' état de traces dans les organismes, le zinc et le fer étant les plus abondants. Les cofacteurs métalliques sont fortement liés à des sites spécifiques des protéines ; bien que les cofacteurs enzymatiques puissent être modifiés au cours de la catalyse, ils retrouvent toujours leur état initial à la fin de la réaction catalysée. Les micronutriments métalliques sont absorbés par les organismes grâce à des transporteurs spécifiques et se lient à des protéines de réserve telles que la ferritine ou la métallothionéine lorsqu'ils ne sont pas utilisés.
Catabolisme
Le processus catabolique le plus courant chez les animaux peut être divisé en trois étapes principales. Dans la première étape, les grosses molécules organiques, telles que les protéines , les polysaccharides ou les lipides , sont digérées en leurs composants plus petits à l'extérieur des cellules. Ensuite, ces molécules plus petites sont absorbées par les cellules et converties en molécules encore plus petites, généralement l'acétyl-coenzyme A (acétyl-CoA), ce qui libère de l'énergie. Enfin, le groupe acétyle de l'acétyl-CoA est oxydé en eau et en dioxyde de carbone au cours du cycle de l'acide citrique et de la chaîne de transport d'électrons , libérant ainsi davantage d'énergie tout en réduisant le nicotinamide adénine dinucléotide (NAD + ) en NADH.
Digestion
Les microbes sécrètent simplement des enzymes digestives dans leur environnement , tandis que les animaux ne sécrètent ces enzymes qu'à partir de cellules spécialisées de leur tube digestif , notamment l' estomac , le pancréas et les glandes salivaires . Les acides aminés ou les sucres libérés par ces enzymes extracellulaires sont ensuite transportés dans les cellules par des protéines de transport actif .
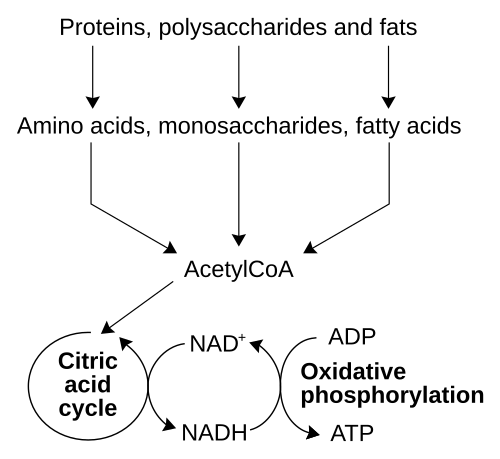
Énergie provenant de composés organiques
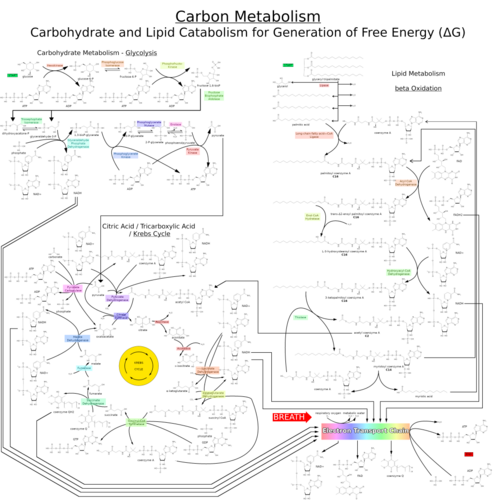
Les lipides sont catabolisés par hydrolyse en acides gras libres et en glycérol. Le glycérol entre dans la glycolyse et les acides gras sont dégradés par β-oxydation pour libérer de l'acétyl-CoA, qui est ensuite intégré au cycle de l'acide citrique. L'oxydation des acides gras libère plus d'énergie que celle des glucides. Les stéroïdes sont également dégradés par certaines bactéries selon un processus similaire à la β-oxydation. Ce processus de dégradation implique la libération de quantités importantes d'acétyl-CoA, de propionyl-CoA et de pyruvate, qui peuvent tous être utilisés par la cellule comme source d'énergie. Mycobacterium tuberculosis peut également se développer en utilisant le cholestérol comme unique source de carbone, et les gènes impliqués dans les voies métaboliques du cholestérol ont été validés comme étant importants à différentes étapes du cycle de vie infectieux de M. tuberculosis .
Les acides aminés sont soit utilisés pour la synthèse des protéines et autres biomolécules, soit oxydés en urée et en dioxyde de carbone pour produire de l'énergie. La voie d'oxydation débute par l'élimination du groupe amine par une transaminase . Ce groupe amine est ensuite intégré au cycle de l'urée , laissant un squelette carboné désaminé sous forme de cétoacide . Plusieurs de ces cétoacides sont des intermédiaires du cycle de l'acide citrique, par exemple l'α- cétoglutarate formé par désamination du glutamate . Les acides aminés glucogéniques peuvent également être convertis en glucose par gluconéogenèse .
Transformations énergétiques
Phosphorylation oxydative
L'expulsion des protons hors des mitochondries crée une différence de concentration protonique de part et d'autre de la membrane et génère un gradient électrochimique . Cette force repousse les protons dans la mitochondrie par la base d'une enzyme appelée ATP synthase . Le flux de protons induit la rotation de la sous-unité de la tige, ce qui modifie la conformation du site actif du domaine synthase et provoque la phosphorylation de l'adénosine diphosphate , la transformant ainsi en ATP.
Énergie provenant de composés inorganiques
L'énergie de la lumière
Chez de nombreux organismes, la capture de l'énergie solaire est similaire, dans son principe, à la phosphorylation oxydative, car elle implique le stockage de l'énergie sous forme de gradient de concentration de protons. Cette force proton-motrice entraîne ensuite la synthèse d'ATP. Les électrons nécessaires au fonctionnement de cette chaîne de transport d'électrons proviennent de protéines photosensibles appelées centres réactionnels photosynthétiques . Ces centres réactionnels sont classés en deux types selon la nature du pigment photosynthétique présent : la plupart des bactéries photosynthétiques n'en possèdent qu'un seul type, tandis que les plantes et les cyanobactéries en possèdent deux.
Chez les plantes, les algues et les cyanobactéries, le photosystème II utilise l'énergie lumineuse pour extraire des électrons de l'eau, libérant ainsi de l'oxygène comme déchet. Ces électrons sont ensuite transférés au complexe cytochrome b6f , qui utilise leur énergie pour pomper des protons à travers la membrane thylakoïde du chloroplaste . Ces protons traversent à nouveau la membrane et activent l'ATP synthase, comme précédemment. Les électrons circulent alors à travers le photosystème I et peuvent être utilisés pour réduire le coenzyme NADP + . Ce coenzyme peut entrer dans le cycle de Calvin ou être recyclé pour la synthèse d'ATP
Anabolisme
L'anabolisme chez les organismes peut varier selon la source des molécules synthétisées dans leurs cellules. Les autotrophes, comme les plantes, peuvent construire des molécules organiques complexes, telles que les polysaccharides et les protéines, à partir de molécules simples comme le dioxyde de carbone et l'eau. Les hétérotrophes , en revanche, nécessitent une source de substances plus complexes, comme les monosaccharides et les acides aminés, pour produire ces molécules complexes. Les organismes peuvent également être classés selon leur source d'énergie : les photoautotrophes et les photohétérotrophes tirent leur énergie de la lumière, tandis que les chimioautotrophes et les chimiohétérotrophes la tirent de réactions d'oxydation.
fixation du carbone
Chez les procaryotes photosynthétiques les mécanismes de fixation du carbone sont plus diversifiés. Le dioxyde de carbone peut y être fixé par le cycle de Calvin-Benson, un cycle de l'acide citrique inversé , ou par la carboxylation de l'acétyl-CoA . Les chimioautotrophes procaryotes fixent également le CO₂ par le cycle de Calvin-Benson, mais utilisent l'énergie de composés inorganiques pour catalyser la réaction
Glucides et glycanes
Bien que les lipides constituent un moyen courant de stockage de l'énergie, chez les vertébrés comme l'homme, les acides gras stockés ne peuvent être convertis en glucose par gluconéogenèse , car ces organismes sont incapables de convertir l'acétyl-CoA en pyruvate ; les plantes, contrairement aux animaux, possèdent l'équipement enzymatique nécessaire. Par conséquent, après un jeûne prolongé, les vertébrés doivent produire des corps cétoniques à partir d'acides gras pour remplacer le glucose dans les tissus, comme le cerveau, qui ne peuvent métaboliser les acides gras. Chez d'autres organismes, tels que les plantes et les bactéries, ce problème métabolique est résolu grâce au cycle du glyoxylate , qui court-circuite l' étape de décarboxylation du cycle de l'acide citrique et permet la transformation de l'acétyl-CoA en oxaloacétate , lequel peut être utilisé pour la production de glucose. Outre les lipides, le glucose est stocké dans la plupart des tissus, constituant une source d'énergie disponible au sein même des tissus grâce à la glycogénèse, qui servait généralement à maintenir la glycémie.
Les polysaccharides et les glycanes sont synthétisés par l'addition séquentielle de monosaccharides, catalysée par une glycosyltransférase , à partir d'un donneur de sucre-phosphate réactif tel que l'uridine diphosphate glucose (UDP-Glc), sur un groupe hydroxyle accepteur du polysaccharide en croissance. Comme n'importe quel groupe hydroxyle du cycle du substrat peut servir d'accepteur, les polysaccharides produits peuvent présenter des structures linéaires ou ramifiées. Ces polysaccharides peuvent avoir des fonctions structurales ou métaboliques intrinsèques, ou être transférés aux lipides et aux protéines par les enzymes oligosaccharyltransférases .
Acides gras, isoprénoïdes et stérols
Les terpènes et les isoprénoïdes constituent une vaste classe de lipides, incluant les caroténoïdes , et forment la plus grande classe de produits naturels végétaux . Ces composés sont synthétisés par l'assemblage et la modification d' unités isoprènes issues des précurseurs réactifs que sont le pyrophosphate d'isopentényle et le pyrophosphate de diméthylallyle . Ces précurseurs peuvent être produits de différentes manières. Chez les animaux et les archées, la voie du mévalonate produit ces composés à partir d'acétyl-CoA , tandis que chez les plantes et les bactéries, la voie non mévalonate utilise le pyruvate et le glycéraldéhyde-3-phosphate comme substrats . Une réaction importante utilisant ces donneurs d'isoprènes activés est la biosynthèse des stérols . Dans ce processus, les unités isoprènes s'assemblent pour former le squalène , puis se replient et s'organisent en cycles pour former le lanostérol . Le lanostérol peut ensuite être converti en d'autres stérols tels que le cholestérol et l'ergostérol .
Protéines
Les acides aminés sont assemblés en protéines par liaisons peptidiques . Chaque protéine possède une séquence unique de résidus d'acides aminés : c'est sa structure primaire . De même que les lettres de l'alphabet peuvent se combiner pour former une variété quasi infinie de mots, les acides aminés peuvent s'assembler selon différentes séquences pour former une grande variété de protéines. Les protéines sont synthétisées à partir d'acides aminés activés par leur fixation à une molécule d'ARN de transfert (ARNt) via une liaison ester . Ce précurseur , l'aminoacyl-ARNt, est produit par une réaction ATP -dépendante catalysée par une aminoacyl-ARNt synthétase . Cet aminoacyl-ARNt sert ensuite de substrat au ribosome , qui fixe l'acide aminé à la chaîne protéique en cours d'élongation, en utilisant l'information de séquence contenue dans un ARN messager .
Synthèse et récupération des nucléotides
xénobiotiques et métabolisme redox
Un problème connexe pour les organismes aérobies est le stress oxydatif . Dans ce contexte, des processus tels que la phosphorylation oxydative et la formation de ponts disulfure lors du repliement des protéines produisent des espèces réactives de l'oxygène , comme le peroxyde d'hydrogène . Ces oxydants nocifs sont éliminés par des métabolites antioxydants tels que le glutathion et des enzymes comme les catalases et les peroxydases .
Thermodynamique des organismes vivants
Réglementation et contrôle
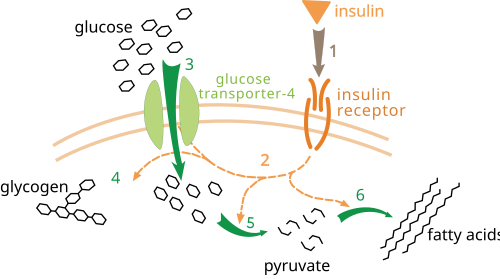
La régulation métabolique comporte plusieurs niveaux. Dans la régulation intrinsèque, la voie métabolique s'autorégule pour répondre aux variations de concentration des substrats ou des produits ; par exemple, une diminution de la quantité de produit peut entraîner une augmentation du flux métabolique pour compenser. Ce type de régulation implique souvent une régulation allostérique de l'activité de plusieurs enzymes de la voie. Le contrôle extrinsèque implique qu'une cellule d'un organisme multicellulaire modifie son métabolisme en réponse à des signaux provenant d'autres cellules. Ces signaux se présentent généralement sous la forme de messagers hydrosolubles, tels que des hormones et des facteurs de croissance , et sont détectés par des récepteurs spécifiques à la surface cellulaire. Ces signaux sont ensuite transmis à l'intérieur de la cellule par des systèmes de seconds messagers , impliquant souvent la phosphorylation de protéines.
Un exemple bien compris de contrôle extrinsèque est la régulation du métabolisme du glucose par l' insuline . L'insuline est produite en réponse à une augmentation de la glycémie . Sa liaison aux récepteurs d'insuline à la surface des cellules active alors une cascade de protéines kinases qui induisent l'absorption du glucose par les cellules et sa conversion en molécules de stockage telles que les acides gras et le glycogène . Le métabolisme du glycogène est contrôlé par l'activité de la phosphorylase , enzyme qui dégrade le glycogène, et de la glycogène synthase , enzyme qui le synthétise. Ces enzymes sont régulées de manière réciproque : la phosphorylation inhibe la glycogène synthase, mais active la phosphorylase. L'insuline induit la synthèse du glycogène en activant les protéines phosphatases et en diminuant la phosphorylation de ces enzymes.
Évolution
De nombreux modèles ont été proposés pour décrire les mécanismes d'évolution des nouvelles voies métaboliques. Parmi ceux-ci figurent l'ajout séquentiel de nouvelles enzymes à une voie ancestrale courte, la duplication puis la divergence de voies entières, ainsi que le recrutement d'enzymes préexistantes et leur assemblage en une nouvelle voie réactionnelle. L'importance relative de ces mécanismes reste incertaine, mais des études génomiques ont montré que les enzymes d'une même voie partagent probablement une origine commune, suggérant que de nombreuses voies ont évolué par étapes, de nouvelles fonctions étant créées à partir d'étapes préexistantes. Un autre modèle, issu d'études retraçant l'évolution des structures protéiques dans les réseaux métaboliques, suggère un recrutement omniprésent des enzymes, qui empruntent des enzymes pour accomplir des fonctions similaires dans différentes voies métaboliques (comme en témoigne la base de données MANET ). Ces processus de recrutement aboutissent à une mosaïque enzymatique évolutive. Une troisième possibilité est que certaines parties du métabolisme puissent exister sous forme de « modules » qui peuvent être réutilisés dans différentes voies et accomplir des fonctions similaires sur différentes molécules.
Outre l’apparition de nouvelles voies métaboliques, l’évolution peut également entraîner la perte de fonctions métaboliques. Par exemple, chez certains parasites , les processus métaboliques non essentiels à la survie disparaissent et les acides aminés, nucléotides et glucides préformés peuvent être récupérés chez l’ hôte . Des capacités métaboliques réduites similaires sont observées chez les organismes endosymbiotiques .
Enquête et manipulation
La figure de droite, illustrant les interactions entre seulement 43 protéines et 40 métabolites, donne un aperçu de la complexité des réseaux métaboliques cellulaires, qui contiennent des milliers d'enzymes différentes : les séquences génomiques fournissent des listes pouvant contenir jusqu'à 26 500 gènes. Il est désormais possible d'utiliser ces données génomiques pour reconstruire des réseaux complets de réactions biochimiques et élaborer des modèles mathématiques plus holistiques , capables d'expliquer et de prédire leur comportement. Ces modèles sont particulièrement performants lorsqu'ils intègrent les données relatives aux voies métaboliques et aux métabolites, obtenues par des méthodes classiques, aux données d' expression génique issues d'études protéomiques et de puces à ADN . Grâce à ces techniques, un modèle du métabolisme humain a été élaboré, qui orientera les futures découvertes de médicaments et la recherche biochimique. Ces modèles sont maintenant utilisés en analyse de réseaux pour classer les maladies humaines en groupes partageant des protéines ou des métabolites communs.
Les réseaux métaboliques bactériens sont un exemple frappant d' organisation en nœud papillon , une architecture capable d'intégrer une large gamme de nutriments et de produire une grande variété de produits et de macromolécules complexes en utilisant relativement peu de devises intermédiaires communes.
Une application technologique majeure de ces informations est l'ingénierie métabolique . Dans ce domaine, des organismes tels que les levures , les plantes ou les bactéries sont génétiquement modifiés afin de les rendre plus utiles en biotechnologie rouge et de faciliter la production de médicaments comme les antibiotiques ou de produits chimiques industriels comme le 1,3-propanediol et l'acide shikimique . Ces modifications génétiques visent généralement à réduire la quantité d'énergie utilisée pour produire le produit, à augmenter les rendements et à réduire la production de déchets.
Histoire
Philosophie grecque
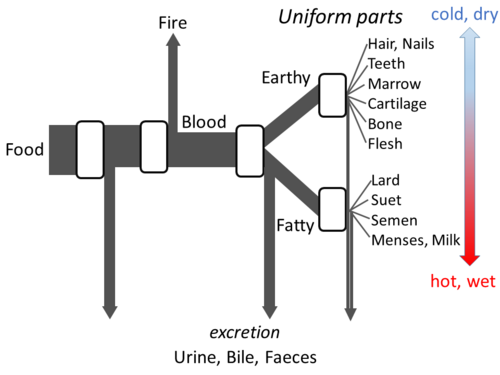
L’ ouvrage d’ Aristote , « Les parties des animaux », expose suffisamment en détail sa conception du métabolisme pour permettre l’élaboration d’un modèle à flux ouvert. Il pensait qu’à chaque étape du processus, les matières issues des aliments étaient transformées, la chaleur étant libérée sous la forme de l’ élément classique du feu, et les matières résiduelles étant excrétées sous forme d’urine, de bile ou de matières fécales.
Ibn al-Nafis a décrit le métabolisme dans son ouvrage de 1260 après J.-C. intitulé Al-Risalah al-Kamiliyyah fil Siera al-Nabawiyyah (Le Traité de Kamil sur la biographie du Prophète) qui comprenait la phrase suivante : « Le corps et ses parties sont dans un état continu de dissolution et de nutrition, ils subissent donc inévitablement un changement permanent. »
Application de la méthode scientifique
L'histoire de l'étude scientifique du métabolisme s'étend sur plusieurs siècles et a évolué, passant de l'examen d'animaux entiers lors des premières études à l'étude des réactions métaboliques individuelles en biochimie moderne. Les premières expériences contrôlées sur le métabolisme humain ont été publiées par Santorio Santorio en 1614 dans son ouvrage *Ars de statica medicina* . Il y décrit comment il se pesait avant et après avoir mangé, dormi , travaillé, eu des rapports sexuels, jeûné, bu et déféqué. Il constata que la majeure partie des aliments ingérés était éliminée par ce qu'il appelait la « transpiration insensible ».

Dans ces premières études, les mécanismes de ces processus métaboliques n'avaient pas été identifiés et l'on pensait qu'une force vitale animait les tissus vivants. Au XIXe siècle, en étudiant la fermentation du sucre en alcool par la levure , Louis Pasteur conclut que la fermentation était catalysée par des substances présentes dans les cellules de levure qu'il nomma « ferments ». Il écrivit que « la fermentation alcoolique est un acte corrélé à la vie et à l'organisation des cellules de levure, et non à leur mort ou à leur putréfaction ». Cette découverte, ainsi que la publication par Friedrich Wöhler en 1828 d'un article sur la synthèse chimique de l' urée , est remarquable car elle représente le premier composé organique préparé à partir de précurseurs entièrement inorganiques. La synthèse de l'urée par Wöhler démontra que des composés organiques pouvaient être créés à partir de précurseurs inorganiques, remettant en question la théorie de la force vitale qui dominait la science du début du XIXe siècle. Les analyses modernes considèrent cette découverte comme fondamentale pour l'unification de la chimie organique et inorganique.
C’est la découverte des enzymes par Eduard Büchner au début du XXe siècle qui a séparé l’étude des réactions chimiques du métabolisme de l’étude biologique des cellules et marqué les débuts de la biochimie . Les connaissances biochimiques ont connu une croissance rapide tout au long du début du XXe siècle. Parmi les biochimistes modernes les plus prolifiques, Hans Krebs a apporté des contributions majeures à l’étude du métabolisme. Il a découvert le cycle de l’urée et, plus tard, en collaboration avec Hans Kornberg , le cycle de l’acide citrique et le cycle du glyoxylate. La recherche biochimique moderne a été grandement facilitée par le développement de nouvelles techniques telles que la chromatographie , la spectroscopie RMN , la microscopie électronique et les simulations de dynamique moléculaire . Ces techniques ont permis la découverte et l’analyse détaillée de nombreuses molécules et voies métaboliques dans les cellules.