Les orchidées sont des plantes appartenant à la famille des Orchidacées ( / ˌ ɔːr k ɪ ˈ d eɪ s i . iː , - s i . aɪ / ), un groupe diversifié et répandu de plantes à fleurs aux fleurs souvent colorées et parfumées. Les orchidées sont des plantes cosmopolites que l'on trouve dans presque tous les habitats de la Terre, à l'exception des glaciers . La plus grande diversité de genres et d'espèces d'orchidées au monde se trouve dans les tropiques .
Les Orchidacées constituent l'une des deux plus grandes familles de plantes à fleurs, avec les Astéracées . Elle comprend environ 28 000 espèces actuellement reconnues, réparties en 763 genres .
La famille des Orchidacées englobe environ 6 à 11 % de toutes les espèces de plantes à graines . Les genres les plus importants sont Bulbophyllum (2 000 espèces), Epidendrum (1 500 espèces), Dendrobium (1 400 espèces) et Pleurothallis (1 000 espèces). Elle comprend également Vanilla (le genre de la plante de vanille ), le genre type Orchis et de nombreuses plantes couramment cultivées telles que Phalaenopsis et Cattleya . De plus, depuis l'introduction des espèces tropicales en culture au XIXe siècle, les horticulteurs ont produit de nombreux hybrides et cultivars .
Description

Les orchidées se distinguent facilement des autres plantes car elles partagent certaines caractéristiques dérivées ou synapomorphies très évidentes . Parmi celles-ci, on trouve : la symétrie bilatérale de la fleur ( zygomorphisme ), de nombreuses fleurs résupinées , un pétale presque toujours très modifié (labelle), des étamines et des carpelles fusionnés et des graines extrêmement petites .
Tige et racines
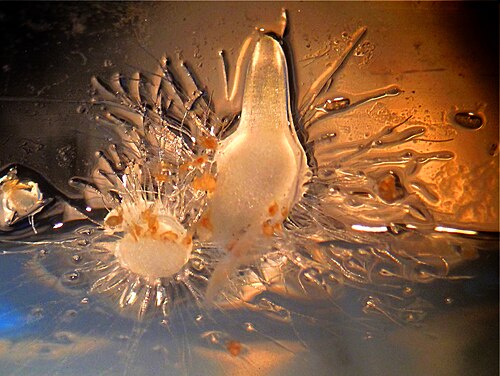
Toutes les orchidées sont des plantes herbacées vivaces dépourvues de toute structure ligneuse permanente . Elles peuvent pousser selon deux schémas :
- Monopodial : La tige pousse à partir d'un seul bourgeon, des feuilles sont ajoutées à partir de l'apex chaque année et la tige s'allonge en conséquence. La tige des orchidées à croissance monopodiale peut atteindre plusieurs mètres de longueur, comme chez Vanda et Vanilla .
- Sympodiale : Les orchidées sympodiales ont une face avant (la croissance la plus récente) et une face arrière (la croissance la plus ancienne). La plante produit une série de pousses adjacentes, qui grandissent jusqu'à une certaine taille, fleurissent puis arrêtent de croître et sont remplacées. Les orchidées sympodiales poussent horizontalement, plutôt que verticalement, en suivant la surface de leur support. La croissance se poursuit par le développement de nouvelles pousses, avec leurs propres feuilles et racines, poussant à partir ou à côté de celles de l'année précédente, comme chez Cattleya . Pendant qu'une nouvelle pousse se développe, le rhizome peut recommencer sa croissance à partir d'un soi-disant « œil », un bourgeon non développé, se ramifiant ainsi. Les orchidées sympodiales peuvent avoir des pseudobulbes visibles reliés par un rhizome , qui rampe le long du dessus ou juste sous le sol.

Les orchidées terrestres peuvent être rhizomateuses ou former des bulbes ou des tubercules . Les coiffes racinaires des orchidées terrestres sont lisses et blanches.
Certaines orchidées terrestres sympodiales, comme Orchis et Ophrys , possèdent deux racines tubéreuses souterraines . L'une sert de réserve alimentaire pour les périodes hivernales, et assure le développement de l'autre, d'où part une croissance visible.
Dans les climats chauds et constamment humides, de nombreuses orchidées terrestres n'ont pas besoin de pseudobulbes.
Les orchidées épiphytes , celles qui poussent sur un support, possèdent des racines aériennes modifiées qui peuvent parfois atteindre plusieurs mètres de long. Dans les parties les plus anciennes des racines, un épiderme spongieux modifié , appelé velamen , a pour fonction d'absorber l'humidité. Il est constitué de cellules mortes et peut avoir une apparence gris argenté, blanche ou brune. Chez certaines orchidées, le velamen comprend des corps spongieux et fibreux à proximité des cellules de passage, appelés tilosomes.
Les cellules de l'épiderme racinaire se développent perpendiculairement à l'axe de la racine pour leur permettre de s'accrocher fermement à leur support. Les nutriments des orchidées épiphytes proviennent principalement de poussières minérales, de détritus organiques, d'excréments d'animaux et d'autres substances qui s'accumulent sur leurs surfaces de support.

La base de la tige des épiphytes sympodiaux, ou chez certaines espèces pratiquement toute la tige, peut être épaissie pour former un pseudobulbe qui contient des nutriments et de l'eau pour les périodes plus sèches.
Le pseudobulbe a généralement une surface lisse avec des rainures longitudinales et peut avoir différentes formes, souvent coniques ou oblongues. Sa taille est très variable ; chez certaines petites espèces de Bulbophyllum , il ne dépasse pas deux millimètres de longueur, alors que chez la plus grande orchidée du monde, Grammatophyllum speciosum (orchidée géante), il peut atteindre trois mètres. Certaines espèces de Dendrobium ont de longs pseudobulbes en forme de canne avec des feuilles courtes et arrondies sur toute la longueur ; d'autres orchidées ont des pseudobulbes cachés ou extrêmement petits, complètement inclus à l'intérieur des feuilles.
En vieillissant, le pseudobulbe perd ses feuilles et entre en dormance. À ce stade, on parle souvent de bulbe arrière. Les bulbes arrière contiennent encore des éléments nutritifs pour la plante, mais ensuite un pseudobulbe prend généralement le relais, exploitant les dernières réserves accumulées dans le bulbe arrière, qui finit lui aussi par mourir. Un pseudobulbe vit généralement environ cinq ans. Les orchidées sans pseudobulbes visibles sont également dites avoir des excroissances, un élément individuel d'une plante sympodiale.
Feuilles
Comme la plupart des monocotylédones , les orchidées ont généralement des feuilles simples à nervures parallèles , bien que certaines Vanilloideae aient une nervure réticulée . Les feuilles peuvent être ovales, lancéolées ou orbiculées et de taille très variable sur chaque plante. Leurs caractéristiques sont souvent diagnostiques. Elles sont normalement alternes sur la tige, souvent pliées dans le sens de la longueur le long du centre (« pliquées »), et n'ont pas de stipules . Les feuilles des orchidées ont souvent des corps siliceux appelés stegmata dans les gaines des faisceaux vasculaires (absents chez les Orchidoideae ) et sont fibreuses.
La structure des feuilles correspond à l'habitat spécifique de la plante. Les espèces qui aiment généralement le soleil ou qui poussent dans des endroits parfois très secs ont des feuilles épaisses et coriaces et les limbes sont recouverts d'une cuticule cireuse pour retenir l'eau nécessaire. Les espèces qui aiment l'ombre, en revanche, ont des feuilles longues et fines.
Les feuilles de la plupart des orchidées sont vivaces, c'est-à-dire qu'elles vivent plusieurs années, tandis que d'autres, en particulier celles aux feuilles plissées comme chez Catasetum , les perdent chaque année et développent de nouvelles feuilles ainsi que de nouveaux pseudobulbes.
Les feuilles de certaines orchidées sont considérées comme ornementales. Les feuilles de Macodes sanderiana , une orchidée semi-terrestre ou rocailleuse (« lithophyte »), présentent des veines argentées et dorées étincelantes sur un fond vert clair. Les feuilles cordées de Psychopsiella limminghei sont vert brun clair avec des marques marron-puce, créées par les pigments des fleurs. La marbrure attrayante des feuilles du sabot de Vénus d'Asie tropicale et subtropicale ( Paphiopedilum ), est due à une répartition inégale de la chlorophylle. De même, Phalaenopsis schilleriana est une orchidée rose pastel aux feuilles tachetées de vert foncé et de vert clair. L'orchidée bijou ( Ludisia discolor ) est davantage cultivée pour ses feuilles colorées que pour ses fleurs blanches.
Certaines orchidées, comme Dendrophylax lindenii (orchidée fantôme), Aphyllorchis et Taeniophyllum, dépendent de leurs racines vertes pour la photosynthèse et n'ont pas de feuilles normalement développées, comme toutes les espèces hétérotrophes .
Les orchidées du genre Corallorhiza (orchidées à racines de corail) n'ont pas du tout de feuilles et ont plutôt des associations symbiotiques ou parasitaires avec le mycélium fongique, à travers lequel elles absorbent les sucres.
Fleurs
Les fleurs d'orchidées ont trois sépales , trois pétales et un ovaire à trois chambres . Les trois sépales et deux des pétales sont souvent similaires les uns aux autres, mais un pétale est généralement très modifié, formant une « lèvre » ou labelle . Dans la plupart des genres d'orchidées, au fur et à mesure que la fleur se développe, elle subit une torsion de 180°, appelée résupination , de sorte que le labelle se trouve sous la colonne . Le labelle a pour fonction d'attirer les insectes et, dans les fleurs résupinées, il sert également de plate-forme d'atterrissage, ou parfois de piège.
.jpg/1280px-Caladenia_alpina_(labelled).jpg)
.jpg/1280px-Diuris_(labelled).jpg)
Les organes reproducteurs d'une fleur d'orchidée sont uniques en ce sens que les étamines et le style sont joints pour former une seule structure, la colonne . Au lieu d'être libérés individuellement, des milliers de grains de pollen sont contenus dans un ou deux faisceaux appelés pollinies qui sont attachés à un disque collant près du sommet de la colonne. Juste en dessous des pollinies se trouve une deuxième plaque collante plus grande appelée stigmate .
Reproduction
Pollinisation
Les mécanismes complexes que les orchidées ont développés pour réaliser la pollinisation croisée ont été étudiés par Charles Darwin et décrits dans Fertilisation of Orchids (1862). Les orchidées ont développé des systèmes de pollinisation hautement spécialisés , de sorte que les chances d'être pollinisées sont souvent rares, de sorte que les fleurs d'orchidées restent généralement réceptives pendant de très longues périodes, ce qui permet aux fleurs non pollinisées de durer longtemps en culture. La plupart des orchidées produisent du pollen en une seule masse. Chaque fois que la pollinisation réussit, des milliers d'ovules peuvent être fécondés.
Les pollinisateurs sont souvent attirés visuellement par la forme et les couleurs du labelle. Cependant, certaines espèces de Bulbophyllum attirent les mouches à fruits mâles ( Bactrocera et Zeugodacus spp.) uniquement par l'intermédiaire d'une substance chimique florale qui agit simultanément comme une récompense florale (par exemple, le méthyleugénol , la cétone de framboise ou la zingérone ) pour effectuer la pollinisation. Les fleurs peuvent produire des odeurs attrayantes. Bien qu'absent chez la plupart des espèces, le nectar peut être produit dans un éperon du labelle ( 8 dans l'illustration ci-dessus), ou sur la pointe des sépales, ou dans les septa de l'ovaire, la position la plus typique chez les Asparagales .

Chez les orchidées qui produisent des pollinies, la pollinisation se déroule selon une variante de la séquence suivante : lorsque le pollinisateur entre dans la fleur, il touche un viscidium, qui se colle aussitôt à son corps, généralement sur la tête ou l'abdomen. En quittant la fleur, il retire le pollinium de l'anthère, car il est relié au viscidium par le caudicule ou le stipe. Le caudicule se courbe alors et le pollinium est déplacé vers l'avant et vers le bas. Lorsque le pollinisateur entre dans une autre fleur de la même espèce, le pollinium a pris une position telle qu'il va se coller au stigmate de la deuxième fleur, juste en dessous du rostellum, la pollinisant. En horticulture, la pollinisation artificielle des orchidées est réalisée en retirant les pollinies avec un petit instrument comme un cure-dent du parent pollen et en les transférant au parent graine.

Certaines orchidées dépendent principalement ou totalement de l'autopollinisation , notamment dans les régions froides où les pollinisateurs sont particulièrement rares. Les caudicules peuvent se dessécher si la fleur n'a été visitée par aucun pollinisateur, et les pollinies tombent alors directement sur le stigmate. Sinon, l'anthère peut tourner et pénétrer alors dans la cavité du stigmate de la fleur (comme chez Holcoglossum amesianum ).
L'orchidée sabot de Vénus Paphiopedilum parishii se reproduit par autofécondation . Cela se produit lorsque l'anthère passe d'un état solide à un état liquide et entre directement en contact avec la surface du stigmate sans l'aide d'un agent pollinisateur ou d'un assemblage floral.
Le labelle des Cypripedioideae est en forme de bonnet et a pour fonction de piéger les insectes visiteurs. La seule sortie mène aux anthères qui déposent le pollen sur le visiteur.
Chez certaines orchidées extrêmement spécialisées, comme le genre eurasien Ophrys , le labelle est adapté pour avoir une couleur, une forme et une odeur qui attirent les insectes mâles par imitation d'une femelle réceptive. La pollinisation se produit lorsque l'insecte tente de s'accoupler avec les fleurs.
De nombreuses orchidées néotropicales sont pollinisées par des abeilles mâles , qui visitent les fleurs pour recueillir des substances chimiques volatiles dont elles ont besoin pour synthétiser des attractifs phéromonaux . On a observé que les mâles d'espèces telles que Euglossa imperialis ou Eulaema meriana quittaient périodiquement leur territoire pour chercher des composés aromatiques, comme le cinéole, afin de synthétiser des phéromones pour attirer et s'accoupler avec les femelles. Chaque type d'orchidée place les pollinies sur une partie différente du corps d'une espèce d'abeille différente, afin de garantir une pollinisation croisée appropriée.
Une orchidée saprophyte achlorophylle rare poussant entièrement sous terre en Australie, Rhizanthella slateri , n'est jamais exposée à la lumière et dépend des fourmis et d'autres insectes terrestres pour la polliniser.
Catasetum , un genre brièvement évoqué par Darwin , lance en réalité ses pollinies visqueuses avec une force explosive lorsqu'un insecte touche une soie , faisant tomber le pollinisateur de la fleur.
Après la pollinisation, les sépales et les pétales se fanent et se flétrissent, mais ils restent généralement attachés à l'ovaire.
En 2011, on a découvert que Bulbophyllum nocturnum fleurissait la nuit.
Reproduction asexuée
Certaines espèces, comme celles des genres Phalaenopsis , Dendrobium et Vanda , produisent des rejets ou des plantules formés à partir d'un des nœuds le long de la tige , grâce à l'accumulation d'hormones de croissance à cet endroit. Ces pousses sont appelées keiki .
Epipogium aphyllum présente une double stratégie de reproduction, s'engageant dans la production de graines à la fois sexuée et asexuée. La probabilité que l'apomixie joue un rôle substantiel dans la réussite de la reproduction semble minime. Dans certains groupes d'espèces de petites orchidées, il existe une préparation notable des gamètes femelles pour la fécondation précédant l'acte de pollinisation.
Fruits et graines

L'ovaire se développe généralement en une capsule déhiscente par trois ou six fentes longitudinales, tout en restant fermée aux deux extrémités .
Les graines sont généralement presque microscopiques et très nombreuses, chez certaines espèces plus d'un million par capsule. Après mûrissement, elles s'envolent comme des particules de poussière ou des spores. La plupart des espèces d'orchidées n'ont pas d'endosperme dans leurs graines et doivent entrer en symbiose avec divers champignons basidiomycètes mycorhiziens qui leur fournissent les nutriments nécessaires à la germination. Ainsi, presque toutes les espèces d'orchidées sont mycohétérotrophes pendant la germination et dépendent des champignons pour compléter leur cycle de vie. Seule une poignée d'espèces d'orchidées ont des graines qui peuvent germer sans mycorhize , à savoir les espèces du genre Disa à graines hydrochorées .

Comme la probabilité qu'une graine rencontre un champignon approprié est très faible, seule une infime partie de toutes les graines libérées se transforment en plantes adultes. En culture, la germination prend généralement des semaines.
Des techniques horticoles ont été mises au point pour faire germer les graines d'orchidées sur un milieu nutritif artificiel, éliminant ainsi le besoin du champignon pour la germination et facilitant grandement la propagation des orchidées ornementales. Le milieu habituel pour le semis des orchidées dans des conditions artificielles est le gel d'agar combiné à une source d'énergie glucidique . La source de glucides peut être une combinaison de sucres discrets ou peut être dérivée d'autres sources telles que la banane , l'ananas , la pêche , voire la purée de tomate ou l'eau de coco . Après la préparation du milieu d'agar, il est versé dans des tubes à essai ou des bocaux qui sont ensuite autoclavés (ou cuits dans un autocuiseur) pour stériliser le milieu. Après la cuisson, le milieu commence à gélifier en refroidissant.
Taxonomie
La taxonomie de cette famille est en constante évolution, car de nouvelles études continuent de clarifier les relations entre les espèces et les groupes d'espèces, permettant de reconnaître davantage de taxons à plusieurs rangs . Les Orchidaceae sont actuellement placées dans l' ordre des Asparagales par le système APG III de 2009.
Cinq sous-familles sont reconnues. Le cladogramme ci-dessous a été établi selon le système APG de 1998. Il représente l'opinion que la plupart des botanistes avaient jusqu'alors défendue. Il a été confirmé par des études morphologiques , mais n'a jamais reçu de soutien solide dans les études de phylogénétique moléculaire .

















.jpg/1280px-Cattleya_Pernell_George_Barnett_"Yankee_Clipper"_(3072486817).jpg)
