Le système du complément , également appelé cascade du complément , fait partie du système immunitaire inné humoral et renforce (complète) la capacité des anticorps et des cellules phagocytaires à éliminer les microbes et les cellules endommagées d'un organisme, à induire une inflammation et à attaquer la membrane cellulaire du pathogène . Bien qu'il appartienne au système immunitaire inné, le système du complément peut être recruté et activé par des anticorps produits par le système immunitaire adaptatif .
Le système du complément est constitué de nombreux précurseurs protéiques inactifs, synthétisés par le foie et circulant dans le sang . Sous l'effet de divers déclencheurs, des protéases du système clivent des protéines spécifiques, libérant ainsi des cytokines et initiant une cascade d'amplification par clivage. Cette cascade d'activation ou de fixation du complément aboutit à la stimulation des phagocytes pour éliminer les substances étrangères et endommagées, à une inflammation attirant davantage de phagocytes et à l'activation du complexe d'attaque membranaire (CAM) , responsable de la destruction des cellules . Le système du complément comprend une cinquantaine de protéines et de fragments protéiques, dont des protéines plasmatiques et des récepteurs membranaires . Ils représentent environ 10 % de la fraction globuline du sérum sanguin.
Trois voies biochimiques activent le système du complément : la voie classique du complément , la voie alterne du complément et la voie des lectines . La voie alterne est responsable de la majorité des activations terminales et, par conséquent, les efforts thérapeutiques dans le traitement des maladies se sont concentrés sur son inhibition.
Histoire
En 1888, George Nuttall découvrit que le sérum sanguin de mouton possédait une faible activité bactéricide contre la bactérie responsable de la maladie du charbon . Cette activité disparaissait lorsqu'il chauffait le sang. En 1891, Hans Ernst August Buchner , observant la même propriété du sang lors de ses expériences, la nomma « alexine », qui signifie « repousser » en grec. Dès 1894, plusieurs laboratoires démontrèrent que le sérum de cobayes guéris du choléra tuait la bactérie du choléra in vitro . Le chauffage du sérum détruisait son activité bactéricide. Néanmoins, le sérum inactivé par la chaleur, injecté à des cobayes exposés à la bactérie du choléra, conservait sa capacité à protéger les animaux contre la maladie. Jules Bordet , un jeune scientifique belge travaillant à l' Institut Pasteur à Paris , a conclu que ce principe comporte deux composantes : l'une conservant un effet sensibilisant après chauffage et l'autre (l'alexine) dont l'effet toxique disparaît après chauffage. La composante thermostable est responsable de l'immunité contre des micro-organismes spécifiques, tandis que la composante thermosensible est responsable de l'activité antimicrobienne non spécifique conférée par tous les sérums normaux. En 1899, Paul Ehrlich a renommé la composante thermosensible « complément ».
Ehrlich a introduit le terme « complément » dans le cadre de sa théorie plus large du système immunitaire. Selon cette théorie, le système immunitaire est constitué de cellules possédant à leur surface des récepteurs spécifiques capables de reconnaître les antigènes . Lors d'une immunisation par un antigène , de nouveaux récepteurs se forment, puis sont libérés par les cellules et circulent dans le sang. Ces récepteurs , que nous appelons aujourd'hui « anticorps », étaient nommés « ambocepteurs » par Ehrlich afin de souligner leur double capacité de liaison : ils reconnaissent et se lient à un antigène spécifique, mais ils reconnaissent et se lient également au composant antimicrobien thermolabile du sérum frais. Ehrlich a donc nommé ce composant thermolabile « complément », car il s'agit d'une substance sanguine qui « complémente » les cellules du système immunitaire. Ehrlich pensait que chaque ambocepteur spécifique d'un antigène possède son propre complément spécifique, tandis que Bordet pensait qu'il n'existe qu'un seul type de complément. Au début du XXe siècle, cette controverse a été résolue lorsqu'il est devenu évident que le complément pouvait agir en combinaison avec des anticorps spécifiques, ou seul de manière non spécifique.
Fonctions

Le complément déclenche les fonctions immunitaires suivantes :
- Attaque membranaire – par rupture de la paroi cellulaire des bactéries ( voie classique du complément )
- Phagocytose – par opsonisation des antigènes. Le C3b possède l'activité opsonisante la plus importante ( voie alterne du complément )
- Inflammation – par attraction des macrophages et des neutrophiles ( voie des lectines )
Aperçu
La plupart des protéines et glycoprotéines constituant le système du complément sont synthétisées par les hépatocytes . Cependant, des quantités importantes sont également produites par les macrophages tissulaires , les monocytes sanguins et les cellules épithéliales des systèmes génito-urinaire et gastro-intestinal . Les trois voies d'activation génèrent toutes des variants homologues de la protéase C3-convertase . La voie classique du complément nécessite généralement des complexes antigène-anticorps pour son activation (réponse immunitaire spécifique), tandis que la voie alterne peut être activée par l'hydrolyse spontanée du composant C3 du complément , par des substances étrangères, des agents pathogènes ou des cellules endommagées. La voie des lectines liant le mannose peut être activée par l'hydrolyse de C3 ou par des antigènes en l'absence d'anticorps (réponse immunitaire non spécifique). Dans les trois voies, la C3-convertase clive et active le composant C3, créant C3a et C3b, et induit une cascade d'événements de clivage et d'activation supplémentaires. Le C3b se lie à la surface des agents pathogènes, ce qui entraîne une plus grande internalisation par les cellules phagocytaires par opsonisation .
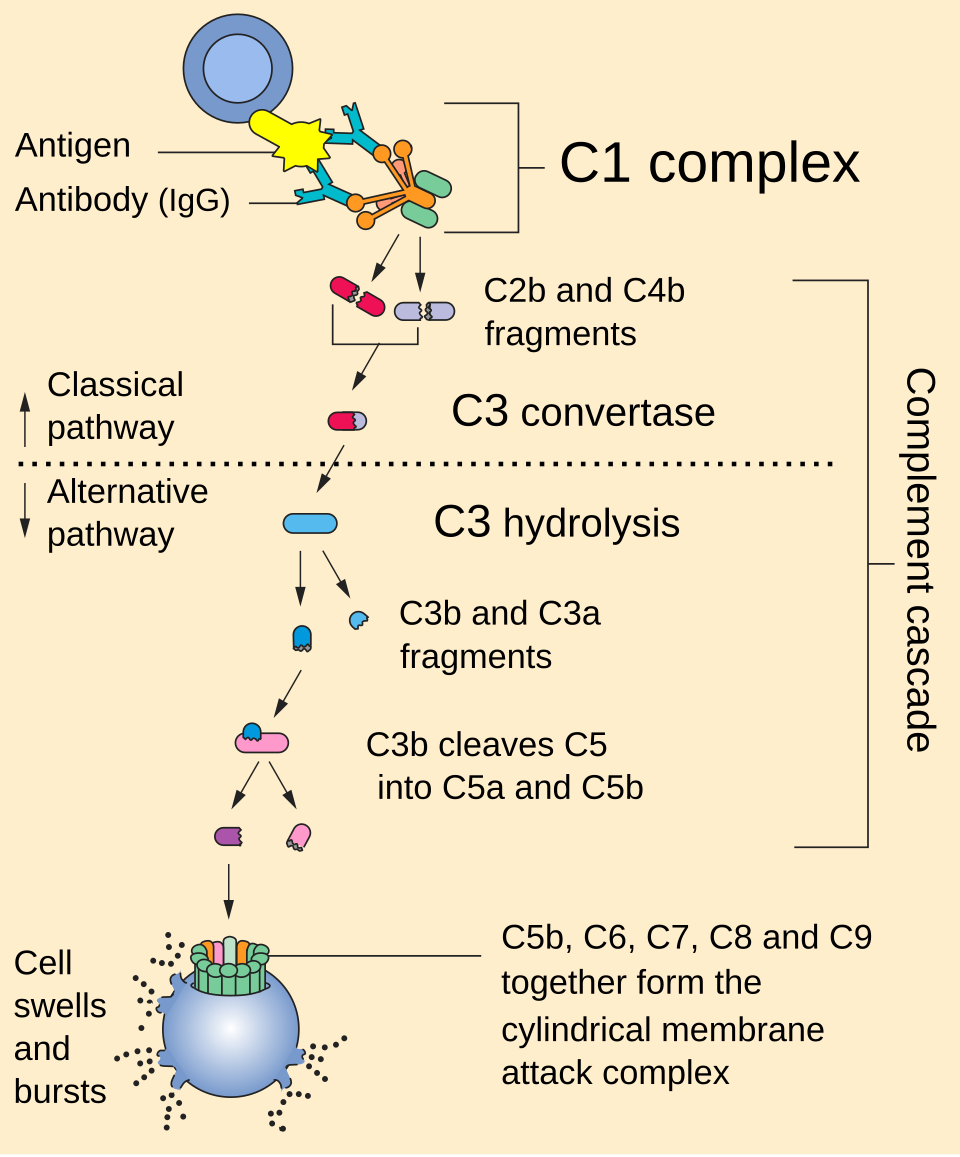
Dans la voie alterne, C3b se lie au facteur B. Le facteur D libère le facteur Ba du facteur B lié à C3b. Le complexe C3b(2)Bb est une protéase qui clive C5 en C5b et C5a. La C5 convertase est également formée par la voie classique lorsque C3b se lie à C4b et C2b. C5a est une protéine chimiotactique importante , contribuant au recrutement des cellules inflammatoires. C3a est le précurseur d'une cytokine importante ( adipokine ) appelée ASP (bien que cette appellation ne soit pas universellement acceptée ) et est généralement clivé rapidement par la carboxypeptidase B. C3a et C5a possèdent tous deux une activité anaphylatoxine , déclenchant directement la dégranulation des mastocytes et augmentant la perméabilité vasculaire et la contraction des muscles lisses . Le C5b initie la voie d'attaque membranaire , aboutissant à la formation du complexe d'attaque membranaire (MAC), composé de C5b, C6 , C7 , C8 et C9 polymérique . Le MAC est le produit final cytolytique de la cascade du complément ; il forme un canal transmembranaire, provoquant la lyse osmotique de la cellule cible. Les cellules de Kupffer et d'autres types de macrophages contribuent à l'élimination des pathogènes recouverts de complément. Intégrés au système immunitaire inné, des éléments de la cascade du complément sont présents chez des espèces antérieures aux vertébrés ; plus récemment, chez la limule, un protostomien , ce qui repousse les origines de ce système à une époque plus reculée qu'on ne le pensait auparavant.

Voie classique
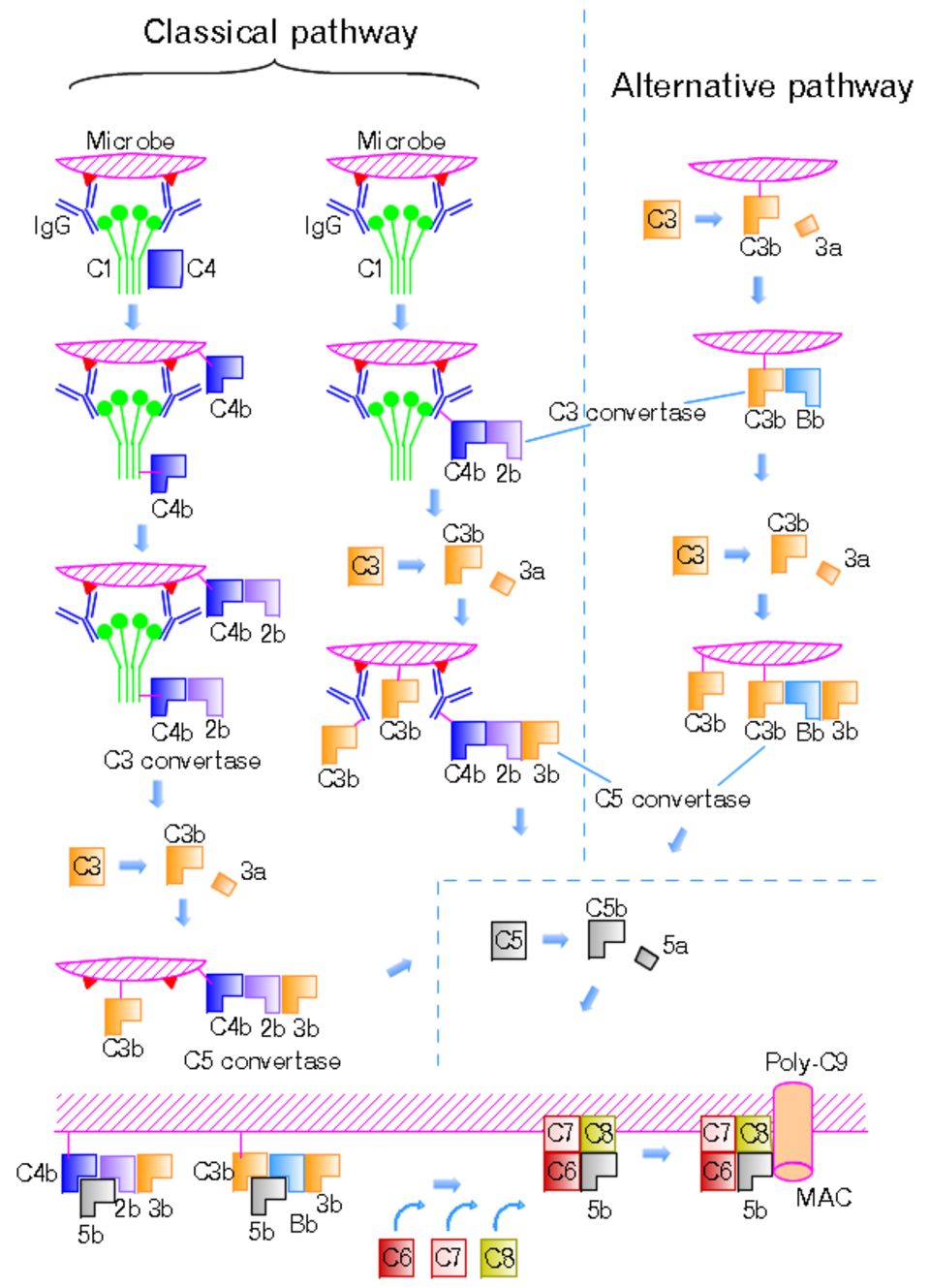
La voie classique est déclenchée par l'activation du complexe C1. Ce complexe est composé d'une molécule de C1q , de deux molécules de C1r et de deux molécules de C1s, soit C1qr₂s₂ . Cette activation a lieu lorsque C1q se lie à un complexe d'IgM ou d'IgG associé à un antigène . Une seule molécule d'IgM pentamérique peut initier la voie, tandis que plusieurs IgG, idéalement six, sont nécessaires. L'activation se produit également lorsque C1q se lie directement à la surface du pathogène. Cette liaison induit des changements conformationnels de la molécule de C1q, ce qui active deux molécules de C1r . C1r est une sérine protéase qui clive ensuite C1s (une autre sérine protéase). Le composant C1r2s2 clive ensuite C4 puis C2 , produisant C4a, C4b, C2a et C2b (historiquement, le plus grand fragment de C2 était appelé C2a , mais il est maintenant désigné par le terme C2b). C4b et C2b s'associent pour former la C3-convertase de la voie classique (complexe C4b2b), qui catalyse le clivage de C3 en C3a et C3b. C3b s'associe ensuite à C4b2b pour former la C5-convertase (complexe C4b2b3b).
Voie alternative
La voie alterne est activée en continu à faible niveau, à l'instar d'un moteur de voiture au ralenti, suite à l'hydrolyse spontanée du C3 due à la rupture de la liaison thioester interne (le C3 est légèrement instable en milieu aqueux). Contrairement aux autres voies, la voie alterne ne dépend pas des anticorps se liant aux pathogènes. Le C3b, généré à partir du C3 par un complexe enzymatique C3 convertase en phase liquide, est rapidement inactivé par les facteurs H et I , de même que le C3 de type C3b, produit du clivage spontané du thioester interne. En revanche, lorsque le thioester interne du C3 réagit avec un groupe hydroxyle ou amine d'une molécule à la surface d'une cellule ou d'un pathogène, le C3b, désormais lié de manière covalente à la surface, est protégé de l'inactivation par le facteur H. Le C3b lié à la surface peut alors se lier au facteur B pour former le C3bB. Ce complexe, en présence du facteur D, sera clivé en Ba et Bb. Bb restera associé à C3b pour former C3bBb, qui est la C3 convertase de la voie alternative.
Le complexe C3bBb est stabilisé par la liaison d'oligomères du facteur P (properdine). La C3 convertase stabilisée, C3bBbP, clive alors une quantité importante de C3, dont une partie se lie de manière covalente à la même surface que C3b. Ce C3b nouvellement lié recrute davantage d'activité des facteurs B, D et P et amplifie considérablement l'activation du complément. Lorsque le complément est activé à la surface d'une cellule, cette activation est limitée par des protéines régulatrices du complément endogènes, notamment CD35 , CD46 , CD55 et CD59 , selon le type cellulaire. Les agents pathogènes, en général, ne possèdent pas de protéines régulatrices du complément (il existe de nombreuses exceptions, qui reflètent l'adaptation des agents pathogènes microbiens aux défenses immunitaires des vertébrés). Ainsi, la voie alterne du complément est capable de distinguer le soi du non-soi en fonction de l'expression des protéines régulatrices du complément à la surface cellulaire. Les cellules hôtes n'accumulent pas de C3b à leur surface (ni son fragment protéolytique appelé iC3b) car ce processus est empêché par les protéines régulatrices du complément, tandis que les cellules étrangères, les pathogènes et les surfaces anormales peuvent être fortement marquées par le C3b et l'iC3b. Par conséquent, la voie alterne du complément constitue un élément de l'immunité innée .
Une fois formée à la surface d'un pathogène ou d'une cellule, l'enzyme C3 convertase alternative peut se lier de manière covalente à une autre molécule de C3b pour former C3bBbC3bP, la C5 convertase. Cette enzyme clive ensuite C5 en C5a, une anaphylatoxine puissante , et en C5b. Ce dernier recrute et assemble alors C6, C7, C8 et plusieurs molécules de C9 pour former le complexe d'attaque membranaire . Ceci crée un pore dans la membrane, capable de tuer ou d'endommager le pathogène ou la cellule.
Voie des lectines
La voie des lectines est homologue à la voie classique, mais utilise l'opsonine, la lectine liant le mannose (MBL), et les ficolines au lieu de C1q. Cette voie est activée par la liaison de la MBL aux résidus mannose à la surface du pathogène, ce qui active les sérine protéases associées à la MBL, MASP-1 et MASP-2 (très similaires à C1r et C1s , respectivement). Ces dernières peuvent alors cliver C4 en C4a et C4b , et C2 en C2a et C2b . C4b et C2b s'associent ensuite pour former la C3-convertase classique , comme dans la voie classique. Les ficolines sont homologues à la MBL et fonctionnent via MASP de manière similaire. Plusieurs polymorphismes nucléotidiques ont été décrits dans le gène de la M-ficoline chez l'homme, influençant l'affinité pour le ligand et les concentrations sériques. Historiquement, le fragment le plus long de C2 était nommé C2a, mais il est maintenant désigné par le nom C2b. Chez les invertébrés dépourvus de système immunitaire adaptatif, les ficolines sont développées et leurs spécificités de liaison diversifiées pour compenser le manque de molécules de reconnaissance spécifiques aux pathogènes.
Nomenclature des fragments de protéines du complément
Les manuels d'immunologie ont utilisé différentes nomenclatures pour les fragments C2 et C2b. Il semble que la désignation privilégiée soit C2a pour le fragment le plus petit : dès 1994, un manuel de référence recommandait de désigner le fragment C2 le plus grand par C2b . Cependant, cette recommandation a été précisée dans la 4e édition de 1999 : « Il est également utile de noter que le fragment actif le plus grand de C2 était initialement désigné C2a, et qu'il l'est encore dans certains ouvrages et articles scientifiques. Par souci de cohérence, nous désignerons ici tous les grands fragments du complément par b , de sorte que le fragment C2 le plus grand sera désigné C2b. Dans les voies classique et des lectines, l'enzyme C3 convertase est formée à partir de C4b membranaire et de C2b. »
Cette nomenclature est également utilisée dans d'autres publications : L'attribution des fragments y est toutefois variable. Certaines sources désignent respectivement les fragments les plus grands et les plus petits par C2a et C2b , tandis que d'autres procèdent inversement Cependant, par convention, C2b désigne ici le fragment le plus grand, qui, selon la méthode classique, forme C4b2b (classiquement C4b2a). Il est à noter que, dans les sept premières éditions de l'ouvrage de Janeway, la dernière abandonne la désignation du fragment C2 comme C2b.
Inhibition virale
Il a également été démontré que la fixation de la protéine MBL sur les surfaces virales améliore la neutralisation des agents pathogènes viraux.
Revoir
| Voie d'activation | Classique | Alternative | Lectine |
|---|---|---|---|
| Activateur | Complexe Ag–Ab | hydrolyse spontanée du C3 | Complexe MBL-mannose |
| C3-convertase | C4b2b | C3bBb | C4b2b |
| C5-convertase | C4b2b3b | C3bBbC3b | C4b2b3b |
| Développement MAC | C5b+C6+C7+C8+C9 | ||
Activation du complément par un anticorps associé à un antigène
Dans la voie classique, C1 se lie, via ses sous-unités C1q, aux fragments Fc (composés de la région CH2) des IgG ou IgM ayant formé un complexe avec les antigènes. C4b et C3b sont également capables de se lier aux IgG ou IgM associées à l'antigène, à leur portion Fc.
Cette liaison du complément, médiée par les immunoglobulines, peut s'interpréter comme suit : le complément utilise la capacité des immunoglobulines à détecter et à se lier aux antigènes non-soi comme point d'ancrage. Le complément lui-même peut se lier aux pathogènes non-soi après avoir détecté leurs motifs moléculaires associés aux pathogènes (PAMP) cependant, grâce à la spécificité des anticorps, le complément peut détecter les cibles non-soi de manière beaucoup plus spécifique.
Certains composants possèdent divers sites de liaison. Dans la voie classique, C4 se lie à C1q associé aux Ig et à l'enzyme C1r2s2, qui clive C4 en C4b et C4a. C4b se lie à C1q, aux Ig associées à l'antigène (plus précisément à leur fragment Fc), et même à la surface du micro-organisme. C3b se lie aux Ig associées à l'antigène et à la surface du micro-organisme. La capacité de C3b à se lier aux Ig associées à l'antigène permettrait de dissocier efficacement les complexes antigène-anticorps.
Règlement
Le système du complément peut être extrêmement nocif pour les tissus de l'hôte, ce qui implique une régulation stricte de son activation. Ce système est régulé par des protéines de contrôle du complément , présentes dans le plasma sanguin et sur la membrane des cellules hôtes . Certaines de ces protéines sont présentes sur les membranes des cellules de l'organisme, les empêchant ainsi d'être ciblées par le complément. C'est le cas, par exemple, de CD59 , également appelée protectine, qui inhibe la polymérisation de C9 lors de la formation du complexe d'attaque membranaire . La voie classique est inhibée par l'inhibiteur de C1 , qui se lie à C1 pour empêcher son activation . Un autre exemple est le facteur H (FH), une protéine plasmatique qui joue un rôle clé dans la régulation négative de la voie alterne . Le facteur H, associé au facteur I , inactive C3b, la forme active de C3. Ce processus empêche la formation de la C3 convertase et interrompt la progression de la cascade du complément. La C3-convertase peut également être inhibée par le facteur d'accélération de la dégradation (DAF), qui est lié aux membranes plasmiques des érythrocytes via une ancre GPI .
Rôle dans la maladie
Déficit du complément
On pense que le système du complément pourrait jouer un rôle dans de nombreuses maladies à composante immunitaire, telles que le syndrome de Barraquer-Simons , l'asthme , le lupus érythémateux , la glomérulonéphrite , diverses formes d' arthrite , les maladies cardiaques auto-immunes , la sclérose en plaques , les maladies inflammatoires de l'intestin , l'hémoglobinurie paroxystique nocturne , le syndrome hémolytique et urémique atypique et les lésions d'ischémie-reperfusion, et le rejet d'organes transplantés.
Il est suggéré que la régulation du complément joue un rôle pendant la grossesse. Une activation inappropriée de la voie alterne du complément pourrait être à l'origine de fausses couches à répétition d'origine immunitaire.
Le système du complément est également de plus en plus impliqué dans les maladies du système nerveux central telles que la maladie d'Alzheimer et d'autres affections neurodégénératives telles que les lésions de la moelle épinière.
Les déficiences de la voie terminale prédisposent à la fois aux maladies auto-immunes et aux infections (en particulier à Neisseria meningitidis , en raison du rôle que joue le complexe d'attaque membranaire (« MAC ») dans l'attaque des bactéries Gram négatives ).
Les infections à N. meningitidis et N. gonorrhoeae sont les seules affections connues pour être associées à des déficits des composants MAC du complément. 40 à 50 % des personnes présentant des déficits MAC souffrent d’infections récurrentes à N. meningitidis .
Déficits en régulateurs du complément
Des mutations des gènes des régulateurs du complément, notamment du facteur H , ont été associées au syndrome hémolytique et urémique atypique . On sont suractivation du complément, soit à la surface des cellules hôtes, soit dans le plasma, la localisation moléculaire des variations génétiques des protéines du complément fournissant des indications sur les mécanismes pathologiques sous-jacents [4]. nucléotidiques et mutations du gène du facteur H du complément (dont la plus fréquente entraîne la substitution p.Y402H) ont été associés à la dégénérescence maculaire liée à l'âge (DMLA ), une maladie oculaire courante . Les polymorphismes du composant C3 du complément , du facteur B du complément et du facteur I du complément, ainsi que la délétion des gènes CfH3 et CfH1, influencent également le risque de développer DMLA .
Les mutations du gène de l'inhibiteur C1 peuvent provoquer un angio-œdème héréditaire , une affection génétique résultant d'une régulation réduite de la bradykinine par le C1-INH.
L’hémoglobinurie paroxystique nocturne est causée par la destruction des globules rouges par le complément en raison d’une incapacité à produire du GPI. Ainsi, les globules rouges ne sont pas protégés par des protéines ancrées au GPI telles que le DAF.
Outils de diagnostic
Les outils diagnostiques permettant de mesurer l'activité du complément comprennent le test d'activité totale du complément .
La présence ou l'absence de fixation du complément après une stimulation peut indiquer la présence d'antigènes ou d'anticorps spécifiques dans le sang. C'est le principe du test de fixation du complément .
Modulation de l'organisme par le complément en cas d'infection
Une activité excessive du système du complément contribue à l'aggravation des symptômes et de la maladie liée à la Covid-19. Bien que le système du complément soit destiné à protéger l'organisme, en situation de stress, il peut engendrer plus de dommages que de protection. Des recherches suggèrent que le système du complément est perturbé lors d'une infection par le VIH / SIDA , ce qui aggrave les lésions corporelles.
Rôle dans le cerveau
Les recherches menées au cours de la dernière décennie ont montré que les protéines du complément de la voie classique du complément jouent un rôle important dans l' élagage synaptique du cerveau au cours du développement précoce.