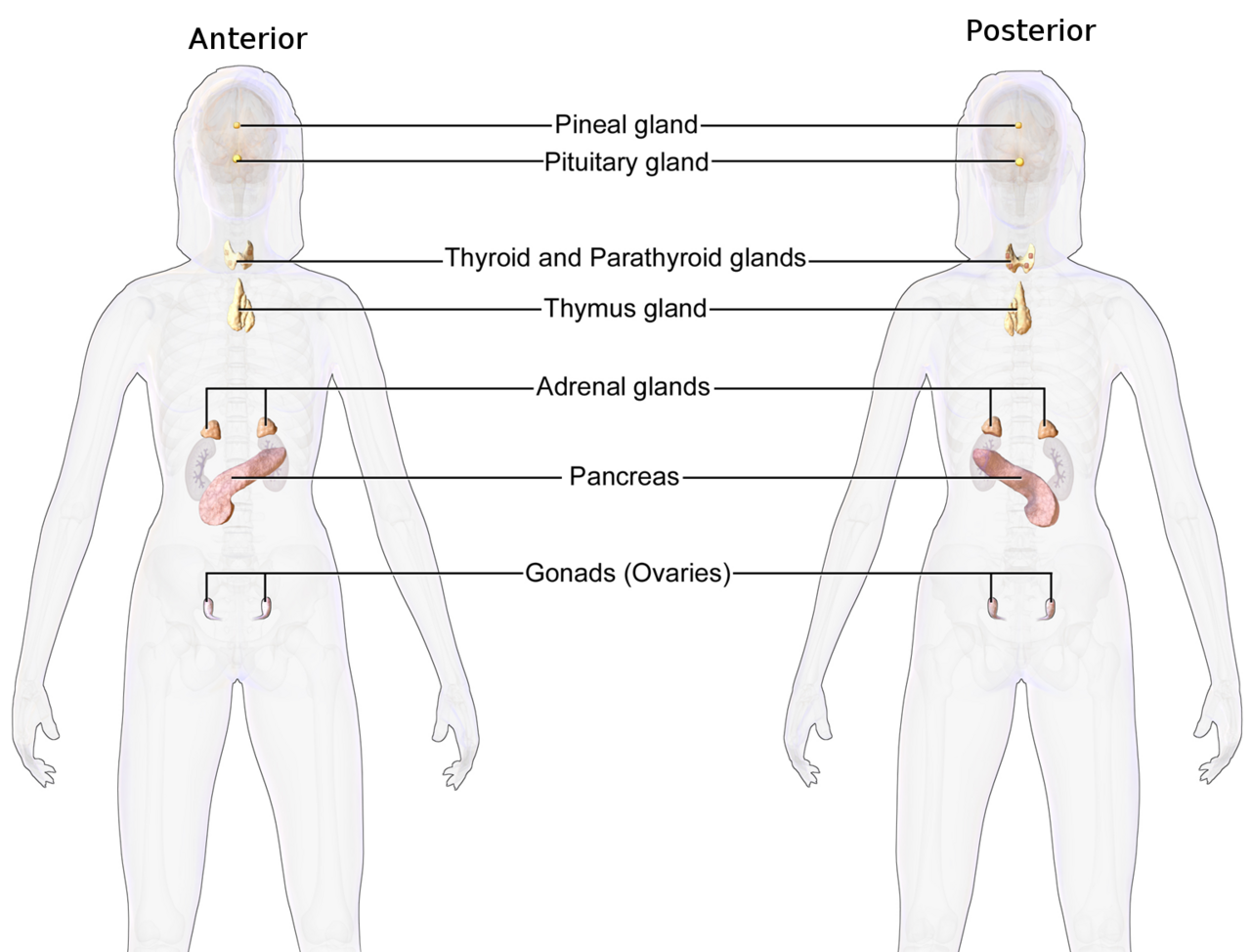
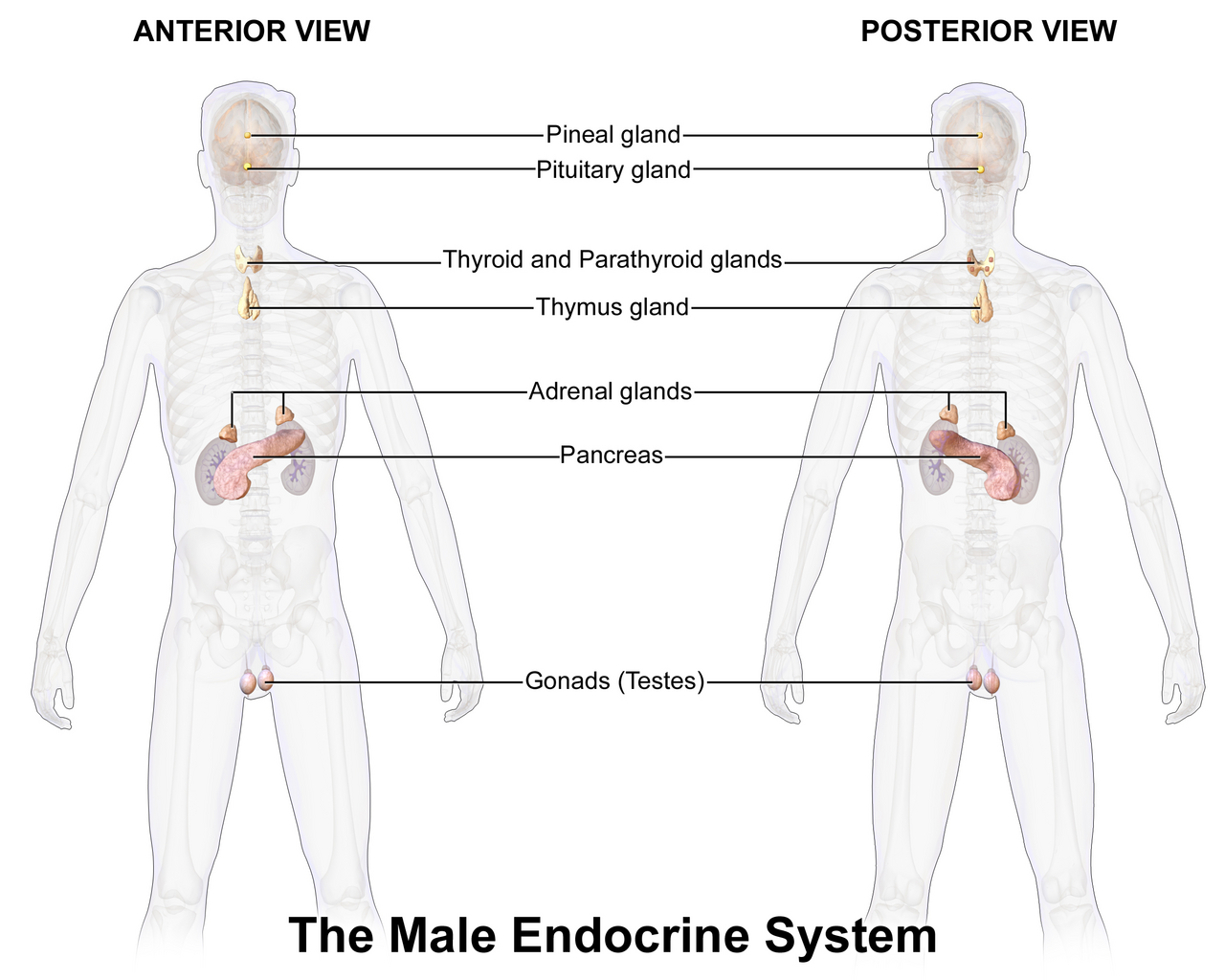
Le système endocrinien est un système de messagerie dans un organisme comprenant des boucles de rétroaction d' hormones qui sont libérées par des glandes internes directement dans le système circulatoire et qui ciblent et régulent des organes distants . Chez les vertébrés , l' hypothalamus est le centre de contrôle neuronal de tous les systèmes endocriniens.
Chez l'homme , les principales glandes endocrines sont la thyroïde , la parathyroïde , l'hypophyse , la glande pinéale et les glandes surrénales , ainsi que les testicules (chez l'homme) et les ovaires (chez la femme) . L' hypothalamus , le pancréas et le thymus font également office de glandes endocrines, entre autres fonctions. (L'hypothalamus et l'hypophyse sont des organes du système neuroendocrinien . L'une des fonctions les plus importantes de l'hypothalamus, situé dans le cerveau à côté de l'hypophyse, est de relier le système endocrinien au système nerveux via l'hypophyse.) D'autres organes, comme les reins , jouent également un rôle au sein du système endocrinien en sécrétant certaines hormones. L'étude du système endocrinien et de ses troubles est connue sous le nom d'endocrinologie . La thyroïde sécrète la thyroxine , l' hypophyse sécrète l'hormone de croissance , la glande pinéale sécrète la mélatonine , les testicules sécrètent la testostérone et les ovaires sécrètent l'œstrogène et la progestérone .
Les glandes qui se signalent les unes les autres en séquence sont souvent appelées un axe, comme l' axe hypothalamo-hypophyso-surrénalien . En plus des organes endocriniens spécialisés mentionnés ci-dessus, de nombreux autres organes qui font partie d'autres systèmes corporels ont des fonctions endocrines secondaires, notamment les os , les reins , le foie , le cœur et les gonades . Par exemple, le rein sécrète l'hormone endocrine érythropoïétine . Les hormones peuvent être des complexes d'acides aminés, des stéroïdes , des eicosanoïdes , des leucotriènes ou des prostaglandines .
Le système endocrinien est à la fois opposé aux glandes exocrines , qui sécrètent des hormones à l'extérieur du corps, et au système connu sous le nom de signalisation paracrine entre les cellules sur une distance relativement courte. Les glandes endocrines n'ont pas de canaux , sont vasculaires et ont généralement des vacuoles ou des granules intracellulaires qui stockent leurs hormones. En revanche, les glandes exocrines, telles que les glandes salivaires , les glandes mammaires et les glandes sous-muqueuses du tube digestif , ont tendance à être beaucoup moins vasculaires et ont des canaux ou une lumière creuse . L'endocrinologie est une branche de la médecine interne .
Structure
Principaux systèmes endocriniens
Le système endocrinien humain est constitué de plusieurs systèmes qui fonctionnent via des boucles de rétroaction . Plusieurs systèmes de rétroaction importants sont véhiculés via l' hypothalamus et l'hypophyse.
- TRH – TSH – T3/T4
- GnRH – LH/FSH – hormones sexuelles
- CRH – ACTH – cortisol
- Rénine – angiotensine – aldostérone
- Leptine vs ghréline
Glandes
Les glandes endocrines sont des glandes du système endocrinien qui sécrètent leurs produits, les hormones , directement dans les espaces interstitiels où elles sont absorbées dans le sang plutôt que par un canal. Les principales glandes du système endocrinien comprennent la glande pinéale , l'hypophyse , le pancréas , les ovaires , les testicules , la glande thyroïde , la glande parathyroïde , l'hypothalamus et les glandes surrénales . L'hypothalamus et l'hypophyse sont des organes neuroendocriniens .
L'hypothalamus et l'hypophyse antérieure sont deux des trois glandes endocrines qui jouent un rôle important dans la signalisation cellulaire. Elles font toutes deux partie de l'axe HPA, connu pour jouer un rôle dans la signalisation cellulaire du système nerveux.
Hypothalamus : L'hypothalamus est un régulateur clé du système nerveux autonome. Le système endocrinien possède trois ensembles de sorties endocrines qui comprennent le système magnocellulaire, le système parvocellulaire et l'intervention autonome. Le système magnocellulaire est impliqué dans l'expression de l'ocytocine ou de la vasopressine. Le système parvocellulaire est impliqué dans le contrôle de la sécrétion d'hormones de l'hypophyse antérieure.
Hypophyse antérieure : Le rôle principal de l'hypophyse antérieure est de produire et de sécréter des hormones tropiques . Certains exemples d'hormones tropiques sécrétées par l'hypophyse antérieure comprennent la TSH, l'ACTH, la GH, la LH et la FSH.
Cellules
Il existe de nombreux types de cellules qui composent le système endocrinien et ces cellules constituent généralement des tissus et des organes plus grands qui fonctionnent à l’intérieur et à l’extérieur du système endocrinien.
- Hypothalamus
- Glande pituitaire antérieure
- Glande pinéale
- Glande pituitaire postérieure
- L'hypophyse postérieure est une partie de l'hypophyse. Cet organe ne produit aucune hormone mais stocke et sécrète des hormones telles que l'hormone antidiurétique (ADH) qui est synthétisée par le noyau supraoptique de l'hypothalamus et l'ocytocine qui est synthétisée par le noyau paraventriculaire de l'hypothalamus. L'ADH a pour fonction d'aider le corps à retenir l'eau ; cela est important pour maintenir un équilibre homéostatique entre les solutions sanguines et l'eau. L'ocytocine a pour fonction d'induire des contractions utérines, de stimuler la lactation et de permettre l'éjaculation.
- Glande thyroïde
- Les cellules folliculaires de la glande thyroïde produisent et sécrètent T 3 et T 4 en réponse à des niveaux élevés de TRH , produits par l' hypothalamus , et à des niveaux élevés ultérieurs de TSH , produits par l' hypophyse antérieure , qui régule en outre l' activité métabolique et le taux de toutes les cellules, y compris la croissance cellulaire et la différenciation tissulaire .
- Glande parathyroïde Le système endocrinien peut contrôler toutes les émotions et peut contrôler la température.
- Les cellules épithéliales des glandes parathyroïdes sont richement irriguées par le sang provenant des artères thyroïdiennes inférieures et supérieures et sécrètent l'hormone parathyroïdienne (PTH). La PTH agit sur les os, les reins et le tube digestif pour augmenter la réabsorption du calcium et l'excrétion du phosphate. De plus, la PTH stimule la conversion de la vitamine D en sa variante la plus active, la 1,25-dihydroxyvitamine D 3 , qui stimule davantage l'absorption du calcium dans le tube digestif.
- Glande du thymus
- Glandes surrénales
- Pancréas
- Le pancréas contient près de 1 à 2 millions d'îlots de Langerhans (un tissu constitué de cellules sécrétant des hormones) et d'acini. Les acini sécrètent des enzymes digestives.
- Cellules alpha
- Les cellules alpha du pancréas sécrètent des hormones pour maintenir l'homéostasie de la glycémie. L'insuline est produite et excrétée pour abaisser la glycémie à des niveaux normaux. Le glucagon, une autre hormone produite par les cellules alpha, est sécrété en réponse à une faible glycémie. Le glucagon stimule les réserves de glycogène dans le foie pour libérer du sucre dans la circulation sanguine afin d'élever la glycémie à des niveaux normaux.
- Cellules bêta
- 60 % des cellules présentes dans les îlots de Langerhans sont des cellules bêta. Les cellules bêta sécrètent de l'insuline . Avec le glucagon, l'insuline aide à maintenir les niveaux de glucose dans notre corps. L'insuline diminue le taux de glucose dans le sang (une hormone hypoglycémiante) tandis que le glucagon augmente le taux de glucose dans le sang.
- Cellules Delta
- Cellules F
- Cellules alpha
- Le pancréas contient près de 1 à 2 millions d'îlots de Langerhans (un tissu constitué de cellules sécrétant des hormones) et d'acini. Les acini sécrètent des enzymes digestives.
- Ovaires
- Testicule
Développement
Le système endocrinien fœtal est l’un des premiers systèmes à se développer au cours du développement prénatal .
Glandes surrénales
Le cortex surrénalien fœtal peut être identifié dans les quatre semaines de gestation . Le cortex surrénalien provient de l'épaississement du mésoderme intermédiaire . À cinq ou six semaines de gestation, le mésonéphros se différencie en un tissu appelé crête génitale. La crête génitale produit les cellules stéroïdogènes des gonades et du cortex surrénalien. La médullosurrénale est dérivée des cellules ectodermiques . Les cellules qui deviendront du tissu surrénalien se déplacent rétropéritonéalement vers la partie supérieure du mésonéphros. À sept semaines de gestation, les cellules surrénales sont rejointes par des cellules sympathiques qui proviennent de la crête neurale pour former la médullosurrénale . À la fin de la huitième semaine, les glandes surrénales ont été encapsulées et ont formé un organe distinct au-dessus des reins en développement. À la naissance, les glandes surrénales pèsent environ huit à neuf grammes (deux fois plus que les glandes surrénales adultes) et représentent 0,5 % du poids corporel total. À 25 semaines, la zone corticosurrénalienne adulte se développe et est responsable de la synthèse primaire des stéroïdes au cours des premières semaines postnatales.
Glande thyroïde
La glande thyroïde se développe à partir de deux groupes différents de cellules embryonnaires. Une partie provient de l'épaississement du plancher pharyngien, qui sert de précurseur aux cellules folliculaires productrices de thyroxine ( T4 ) . L'autre partie provient des extensions caudales des quatrièmes poches pharyngobranchiales qui donnent naissance aux cellules parafolliculaires sécrétant la calcitonine. Ces deux structures sont apparentes entre 16 et 17 jours de gestation. Vers le 24e jour de gestation, le foramen caecum , un diverticule fin en forme de flacon de l' ébauche médiane , se développe. Entre 24 et 32 jours de gestation environ, l'ébauche médiane se développe en une structure bilobée. À 50 jours de gestation, les ébauches médiale et latérale ont fusionné. À 12 semaines de gestation, la thyroïde fœtale est capable de stocker de l'iode pour la production de TRH , de TSH et d'hormone thyroïdienne libre. À 20 semaines, le fœtus est capable de mettre en œuvre des mécanismes de rétroaction pour la production d'hormones thyroïdiennes. Au cours du développement fœtal, la T 4 est la principale hormone thyroïdienne produite, tandis que la triiodothyronine (T 3 ) et son dérivé inactif, la T 3 inverse , ne sont détectés qu'au troisième trimestre.
Glandes parathyroïdes
Vue latérale et ventrale d'un embryon montrant les troisième (inférieure) et quatrième (supérieure) glandes parathyroïdes au cours de la 6e semaine d'embryogenèse
Une fois que l'embryon atteint quatre semaines de gestation, les glandes parathyroïdes commencent à se développer. L'embryon humain forme cinq ensembles de poches pharyngiennes tapissées d' endoderme . La troisième et la quatrième poche sont responsables du développement des glandes parathyroïdes inférieures et supérieures, respectivement. La troisième poche pharyngienne rencontre la glande thyroïde en développement et elles migrent vers les pôles inférieurs des lobes thyroïdiens. La quatrième poche pharyngienne rencontre plus tard la glande thyroïde en développement et migre vers les pôles supérieurs des lobes thyroïdiens. À 14 semaines de gestation, les glandes parathyroïdes commencent à s'élargir de 0,1 mm de diamètre à environ 1 à 2 mm à la naissance. Les glandes parathyroïdes en développement sont physiologiquement fonctionnelles à partir du deuxième trimestre.
Des études sur des souris ont montré que l'interférence avec le gène HOX15 peut provoquer une aplasie de la glande parathyroïde , ce qui suggère que le gène joue un rôle important dans le développement de la glande parathyroïde. Il a également été démontré que les gènes TBX1 , CRKL , GATA3 , GCM2 et SOX3 jouent un rôle crucial dans la formation de la glande parathyroïde. Les mutations des gènes TBX1 et CRKL sont corrélées au syndrome de DiGeorge , tandis que les mutations du gène GATA3 ont également entraîné un syndrome de type DiGeorge. Les malformations du gène GCM2 ont entraîné une hypoparathyroïdie . Des études sur les mutations du gène SOX3 ont démontré qu'il joue un rôle dans le développement de la parathyroïde. Ces mutations entraînent également divers degrés d'hypopituitarisme.
Pancréas
Le pancréas fœtal humain commence à se développer à partir de la quatrième semaine de gestation. Cinq semaines plus tard, les cellules alpha et bêta pancréatiques ont commencé à émerger. À l'âge de huit à dix semaines de développement, le pancréas commence à produire de l'insuline , du glucagon , de la somatostatine et du polypeptide pancréatique . Au cours des premiers stades du développement fœtal, le nombre de cellules alpha pancréatiques dépasse celui des cellules bêta pancréatiques . Les cellules alpha atteignent leur maximum au milieu de la gestation. Du milieu de la gestation jusqu'au terme, le nombre de cellules bêta continue d'augmenter jusqu'à atteindre un rapport approximatif de 1:1 avec les cellules alpha. La concentration d'insuline dans le pancréas fœtal est de 3,6 pmol/g à sept à dix semaines, et passe à 30 pmol/g à 16-25 semaines de gestation. À court terme, la concentration d'insuline augmente à 93 pmol/g. Les cellules endocrines se sont dispersées dans tout le corps en 10 semaines. À 31 semaines de développement, les îlots de Langerhans se sont différenciés.
Bien que le pancréas fœtal possède des cellules bêta fonctionnelles entre 14 et 24 semaines de gestation, la quantité d'insuline libérée dans la circulation sanguine est relativement faible. Dans une étude réalisée sur des femmes enceintes portant des fœtus à mi-gestation et à un stade proche du terme de leur développement, les fœtus n'ont pas présenté d'augmentation des taux d'insuline plasmatique en réponse à des injections de taux élevés de glucose. Contrairement à l'insuline, les taux plasmatiques de glucagon fœtal sont relativement élevés et continuent d'augmenter au cours du développement. À mi-gestation, la concentration de glucagon est de 6 μg/g, contre 2 μg/g chez l'adulte. Tout comme l'insuline, les taux plasmatiques de glucagon fœtal ne changent pas en réponse à une perfusion de glucose. Cependant, une étude sur une perfusion d'alanine chez des femmes enceintes a montré qu'elle augmentait les concentrations de glucagon dans le sang du cordon ombilical et chez la mère, démontrant ainsi une réponse fœtale à l'exposition aux acides aminés.
Ainsi, alors que les cellules alpha et bêta des îlots pancréatiques fœtaux sont pleinement développées et sont capables de synthétiser des hormones pendant la maturation fœtale restante, les cellules des îlots sont relativement immatures dans leur capacité à produire du glucagon et de l'insuline. On pense que cela résulte des niveaux relativement stables de concentrations de glucose sérique fœtal obtenus via le transfert maternel de glucose à travers le placenta. D'autre part, les niveaux stables de glucose sérique fœtal pourraient être attribués à l'absence de signalisation pancréatique initiée par les incrétines pendant l'alimentation. De plus, les cellules des îlots pancréatiques fœtaux sont incapables de produire suffisamment d'AMPc et dégradent rapidement l'AMPc par la phosphodiestérase nécessaire à la sécrétion de glucagon et d'insuline.
Au cours du développement fœtal, le stockage du glycogène est contrôlé par les glucocorticoïdes fœtaux et le lactogène placentaire . L'insuline fœtale est responsable de l'augmentation de l'absorption du glucose et de la lipogenèse au cours des étapes précédant la naissance. Les cellules fœtales contiennent une plus grande quantité de récepteurs d'insuline par rapport aux cellules adultes et les récepteurs d'insuline fœtale ne sont pas régulés à la baisse en cas d' hyperinsulinémie . En comparaison, les récepteurs haptiques du glucagon fœtal sont abaissés par rapport aux cellules adultes et l'effet glycémique du glucagon est atténué. Ce changement physiologique temporaire contribue à l'augmentation du taux de développement fœtal au cours du dernier trimestre. Un diabète sucré maternel mal géré est lié à la macrosomie fœtale , à un risque accru de fausse couche et à des anomalies du développement fœtal. L'hyperglycémie maternelle est également liée à une augmentation des taux d'insuline et à une hyperplasie des cellules bêta chez le nourrisson post-terme. Les enfants de mères diabétiques présentent un risque accru de maladies telles que : la polycythémie , la thrombose veineuse rénale , l’hypocalcémie , le syndrome de détresse respiratoire , la jaunisse , la cardiomyopathie , les maladies cardiaques congénitales et le développement anormal des organes.
Gonades
Le développement de l'appareil reproducteur débute entre la quatrième et la cinquième semaine de gestation par la migration des cellules germinales. La gonade bipotentielle résulte de la collection de la région médioventrale de la crête urogénitale . Au bout de cinq semaines, les gonades en développement se détachent du primordium surrénalien. La différenciation gonadique commence 42 jours après la conception.
Développement des gonades mâles
Chez les mâles, les testicules se forment à six semaines de gestation et les cellules de Sertoli commencent à se développer à partir de la huitième semaine de gestation. SRY , le locus déterminant le sexe, sert à différencier les cellules de Sertoli . Les cellules de Sertoli sont le point d'origine de l'hormone anti-Müllerienne . Une fois synthétisée, l'hormone anti-Müllerienne initie la régression ipsilatérale du tractus müllerien et inhibe le développement des caractéristiques internes féminines. À 10 semaines de gestation, les cellules de Leydig commencent à produire des hormones androgènes. L'hormone androgène dihydrotestostérone est responsable du développement des organes génitaux externes masculins.
Les testicules descendent au cours du développement prénatal dans un processus en deux étapes qui commence à la huitième semaine de gestation et se poursuit jusqu'au milieu du troisième trimestre. Au cours de la phase transabdominale (8 à 15 semaines de gestation), le ligament gubernaculaire se contracte et commence à s'épaissir. Le ligament craniosuspenseur commence à se dégrader. Cette étape est régulée par la sécrétion d' insuline-like 3 (INSL3), un facteur de type relaxine produit par les testicules, et du récepteur couplé à INSL3 G, LGR8. Au cours de la phase transinguinale (25 à 35 semaines de gestation), les testicules descendent dans le scrotum. Cette étape est régulée par les androgènes, le nerf génito-fémoral et le peptide lié au gène de la calcitonine. Au cours des deuxième et troisième trimestres, le développement testiculaire se termine par la diminution des cellules de Leydig fœtales et l'allongement et l'enroulement des cordons séminifères .
Développement des gonades féminines
Chez la femme, les ovaires deviennent morphologiquement visibles à partir de la 8e semaine de gestation. L'absence de testostérone entraîne la diminution des structures de Wolff. Les structures de Müller demeurent et se développent en trompes de Fallope, en utérus et dans la partie supérieure du vagin. Le sinus urogénital se développe en urètre et dans la partie inférieure du vagin, le tubercule génital se développe en clitoris, les plis urogénitaux se développent en petites lèvres et les gonflements urogénitaux se développent en grandes lèvres. À 16 semaines de gestation, les ovaires produisent des récepteurs FSH et LH/hCG . À 20 semaines de gestation, les précurseurs des cellules de la thèque sont présents et la mitose des oogones se produit. À 25 semaines de gestation, l'ovaire est morphologiquement défini et la folliculogenèse peut commencer.
Des études sur l'expression génétique montrent qu'un ensemble spécifique de gènes, tels que la follistatine et plusieurs inhibiteurs de la cycline kinase, sont impliqués dans le développement ovarien. Il a été démontré qu'un ensemble de gènes et de protéines, tels que WNT4, RSPO1, FOXL2 et divers récepteurs d'œstrogènes, empêchent le développement des testicules ou la lignée de cellules de type mâle.
Glande pituitaire
L' hypophyse se forme dans la plaque neurale rostrale. La poche de Rathke, une cavité de cellules ectodermiques de l' oropharynx , se forme entre la quatrième et la cinquième semaine de gestation et, une fois complètement développée, donne naissance à l'hypophyse antérieure. À la septième semaine de gestation, le système vasculaire de l'hypophyse antérieure commence à se développer. Au cours des 12 premières semaines de gestation, l'hypophyse antérieure subit une différenciation cellulaire. À la 20e semaine de gestation, le système porte hypophysaire s'est développé. La poche de Rathke se développe vers le troisième ventricule et fusionne avec le diverticule. Cela élimine la lumière et la structure devient la fente de Rathke. Le lobe postérieur de l'hypophyse est formé à partir du diverticule. Des portions du tissu hypophysaire peuvent rester dans la ligne médiane du nasopharynx. Dans de rares cas, cela entraîne des tumeurs ectopiques fonctionnelles sécrétant des hormones dans le nasopharynx.
Le développement fonctionnel de l'hypophyse antérieure implique une régulation spatiotemporelle des facteurs de transcription exprimés dans les cellules souches hypophysaires et des gradients dynamiques de facteurs solubles locaux. La coordination du gradient dorsal de la morphogenèse hypophysaire dépend des signaux neuroectodermiques de la protéine morphogénétique osseuse infundibulaire 4 (BMP4). Cette protéine est responsable du développement de l'invagination initiale de la poche de Rathke. Les autres protéines essentielles nécessaires à la prolifération des cellules hypophysaires sont le facteur de croissance des fibroblastes 8 (FGF8), Wnt4 et Wnt5. Le schéma de développement ventral et l'expression des facteurs de transcription sont influencés par les gradients de BMP2 et de la protéine Sonic Hedgehog (SHH). Ces facteurs sont essentiels à la coordination des schémas précoces de prolifération cellulaire.
Six semaines après le début de la gestation, les cellules corticotropes peuvent être identifiées. À sept semaines de gestation, l'hypophyse antérieure est capable de sécréter de l'ACTH. Dans les huit semaines de gestation, les cellules somatotropes commencent à se développer avec une expression cytoplasmique de l'hormone de croissance humaine. Une fois que le fœtus atteint 12 semaines de développement, les thyrotropes commencent à exprimer les sous-unités bêta de la TSH, tandis que les gonadotropes commencent à exprimer les sous-unités bêta de la LH et de la FSH. Les fœtus mâles produisent principalement des gonadotropes exprimant la LH, tandis que les fœtus femelles produisent une expression égale de gonadotropes exprimant la LH et la FSH. À 24 semaines de gestation, les lactotropes exprimant la prolactine commencent à émerger.
Fonction
Hormones
Une hormone est une molécule de signalisation produite par les cellules des glandes des organismes multicellulaires et transportée par le système circulatoire vers des organes distants pour réguler la physiologie et le comportement . Les hormones ont des structures chimiques diverses, principalement de 3 classes : les eicosanoïdes , les stéroïdes et les dérivés d'acides aminés / protéines ( amines , peptides et protéines ). Les glandes qui sécrètent des hormones constituent le système endocrinien. Le terme hormone est parfois étendu pour inclure les substances chimiques produites par les cellules qui affectent la même cellule ( signalisation autocrine ou intracrine ) ou les cellules voisines ( signalisation paracrine ).
Les hormones sont utilisées pour communiquer entre les organes et les tissus pour la régulation physiologique et les activités comportementales , telles que la digestion, le métabolisme , la respiration , la fonction tissulaire , la perception sensorielle , le sommeil , l'excrétion , la lactation , le stress , la croissance et le développement , le mouvement , la reproduction et l'humeur .
Les hormones affectent les cellules distantes en se liant à des protéines réceptrices spécifiques dans la cellule cible, ce qui entraîne un changement dans la fonction cellulaire. Cela peut conduire à des réponses spécifiques au type de cellule qui incluent des changements rapides de l'activité des protéines existantes ou des changements plus lents dans l' expression des gènes cibles. Les hormones à base d'acides aminés ( amines et hormones peptidiques ou protéiques ) sont hydrosolubles et agissent à la surface des cellules cibles via des voies de transduction du signal ; les hormones stéroïdes , étant liposolubles, se déplacent à travers les membranes plasmiques des cellules cibles pour agir dans leurs noyaux .
Signalisation cellulaire
Le mode de signalisation cellulaire typique dans le système endocrinien est la signalisation endocrine, c'est-à-dire l'utilisation du système circulatoire pour atteindre des organes cibles éloignés. Cependant, il existe également d'autres modes, à savoir la signalisation paracrine, autocrine et neuroendocrine . La signalisation purement neurocrine entre neurones , en revanche, appartient entièrement au système nerveux .
Autocrine
La signalisation autocrine est une forme de signalisation dans laquelle une cellule sécrète une hormone ou un messager chimique (appelé agent autocrine) qui se lie aux récepteurs autocrines de la même cellule, entraînant des changements dans les cellules.
Paracrine
Certains endocrinologues et cliniciens incluent le système paracrine dans le système endocrinien, mais il n'y a pas de consensus. Les paracrines agissent plus lentement et ciblent les cellules du même tissu ou organe. La somatostatine , par exemple , est libérée par certaines cellules pancréatiques et cible d'autres cellules pancréatiques.
Juxtacrine
La signalisation juxtacrine est un type de communication intercellulaire qui est transmise via des composants oligosaccharidiques, lipidiques ou protéiques d'une membrane cellulaire et peut affecter soit la cellule émettrice, soit les cellules immédiatement adjacentes.
Elle se produit entre des cellules adjacentes qui possèdent de larges zones de membrane plasmique étroitement opposées reliées par des canaux transmembranaires appelés connexons . L'écart entre les cellules peut généralement être compris entre 2 et 4 nm seulement.
Importance clinique
Maladie

Les maladies du système endocrinien sont courantes, notamment le diabète sucré , les maladies thyroïdiennes et l'obésité . Les maladies endocriniennes sont caractérisées par une libération hormonale mal régulée ( adénome hypophysaire productif ), une réponse inappropriée à la signalisation ( hypothyroïdie ), l'absence d'une glande ( diabète sucré de type 1 , diminution de l'érythropoïèse en cas d'insuffisance rénale chronique ) ou une hypertrophie structurelle dans un site critique comme la thyroïde ( goitre multinodulaire toxique ). L'hypofonctionnement des glandes endocrines peut survenir à la suite d'une perte de réserve, d'une hyposécrétion, d'une agénésie , d'une atrophie ou d'une destruction active. L'hyperfonctionnement peut survenir à la suite d'une hypersécrétion, d'une perte de suppression, d' un changement hyperplasique ou néoplasique ou d'une hyperstimulation.
Les endocrinopathies sont classées comme primaires, secondaires ou tertiaires. La maladie endocrinienne primaire inhibe l'action des glandes en aval. La maladie endocrinienne secondaire indique un problème avec l'hypophyse. La maladie endocrinienne tertiaire est associée à un dysfonctionnement de l'hypothalamus et de ses hormones libératrices.
Comme la thyroïde et les hormones ont été impliquées dans la signalisation de prolifération de tissus distants, par exemple, il a été démontré que le récepteur d'œstrogène est impliqué dans certains cancers du sein . La signalisation endocrine, paracrine et autocrine a été impliquée dans la prolifération, l'une des étapes nécessaires de l'oncogenèse .
D'autres maladies courantes résultant d'un dysfonctionnement endocrinien comprennent la maladie d'Addison , la maladie de Cushing et la maladie de Graves . La maladie de Cushing et la maladie d'Addison sont des pathologies impliquant le dysfonctionnement de la glande surrénale. Le dysfonctionnement de la glande surrénale peut être dû à des facteurs primaires ou secondaires et peut entraîner un hypercortisolisme ou un hypocortisolisme . La maladie de Cushing est caractérisée par l'hypersécrétion de l'hormone adrénocorticotrope (ACTH) due à un adénome hypophysaire qui provoque finalement un hypercortisolisme endogène en stimulant les glandes surrénales. Certains signes cliniques de la maladie de Cushing comprennent l'obésité, le visage lunaire et l'hirsutisme. La maladie d'Addison est une maladie endocrinienne qui résulte d'un hypocortisolisme causé par une insuffisance des glandes surrénales. L'insuffisance surrénale est importante car elle est corrélée à une diminution de la capacité à maintenir la pression artérielle et la glycémie, un défaut qui peut s'avérer fatal.
La maladie de Graves implique une hyperactivité de la glande thyroïde qui produit les hormones T3 et T4. de la maladie de Graves vont de la transpiration excessive, de la fatigue , de l'intolérance à la chaleur et de l'hypertension artérielle à un gonflement des yeux qui provoque des rougeurs, des gonflements et, dans de rares cas, une vision réduite ou double.
Autres animaux
Un système neuroendocrinien a été observé chez tous les animaux dotés d'un système nerveux et tous les vertébrés ont un axe hypothalamus-hypophyse. Tous les vertébrés ont une thyroïde, qui chez les amphibiens est également cruciale pour la transformation des larves en forme adulte. Tous les vertébrés ont un tissu de glande surrénale, les mammifères étant uniques en ce sens qu'il est organisé en couches. Tous les vertébrés ont une forme d'axe rénine-angiotensine, et tous les tétrapodes ont l'aldostérone comme minéralocorticoïde primaire .
Images supplémentaires
-
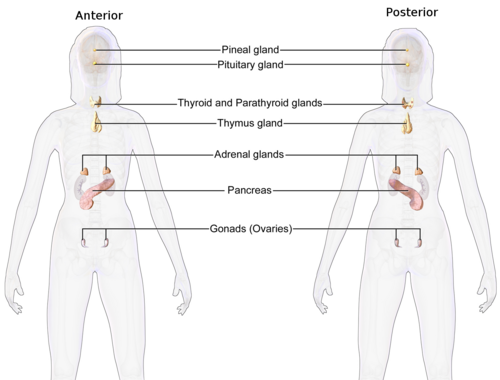 Système endocrinien féminin
Système endocrinien féminin -
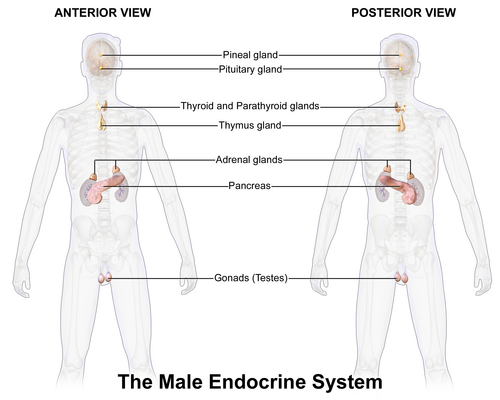 Système endocrinien masculin
Système endocrinien masculin