La protéine du domaine de mort associée au FAS , également appelée MORT1 , est codée par le gène FADD sur la région 11q13.3 du chromosome 11 chez l'homme.
FADD est une protéine adaptatrice qui relie les membres de la superfamille des récepteurs du facteur de nécrose tumorale , tels que le récepteur Fas , aux procaspases 8 et 10 pour former le complexe de signalisation induisant la mort (DISC) pendant l'apoptose . En plus de son rôle le plus connu dans l'apoptose, FADD a également été observé comme jouant un rôle dans d'autres processus, notamment la prolifération, la régulation du cycle cellulaire et le développement.
Structure
FADD est une protéine de 23 kDa, composée de 208 acides aminés. Elle contient deux domaines principaux : un domaine de mort terminal C (DD) et un domaine effecteur de mort terminal N (DED). Chaque domaine, bien que partageant très peu de similarité de séquence, est structurellement similaire l'un à l'autre, chacun étant constitué de 6 hélices α. Le DD de FADD se lie à des récepteurs tels que le récepteur Fas à la membrane plasmique via leur DD. L'interaction entre les domaines de mort est une interaction électrostatique impliquant les hélices α 2 et 3 du domaine à 6 hélices. Le DED se lie au DED de molécules intracellulaires telles que la procaspase 8. On pense que cette interaction se produit par le biais d'interactions hydrophobes .
Fonctions
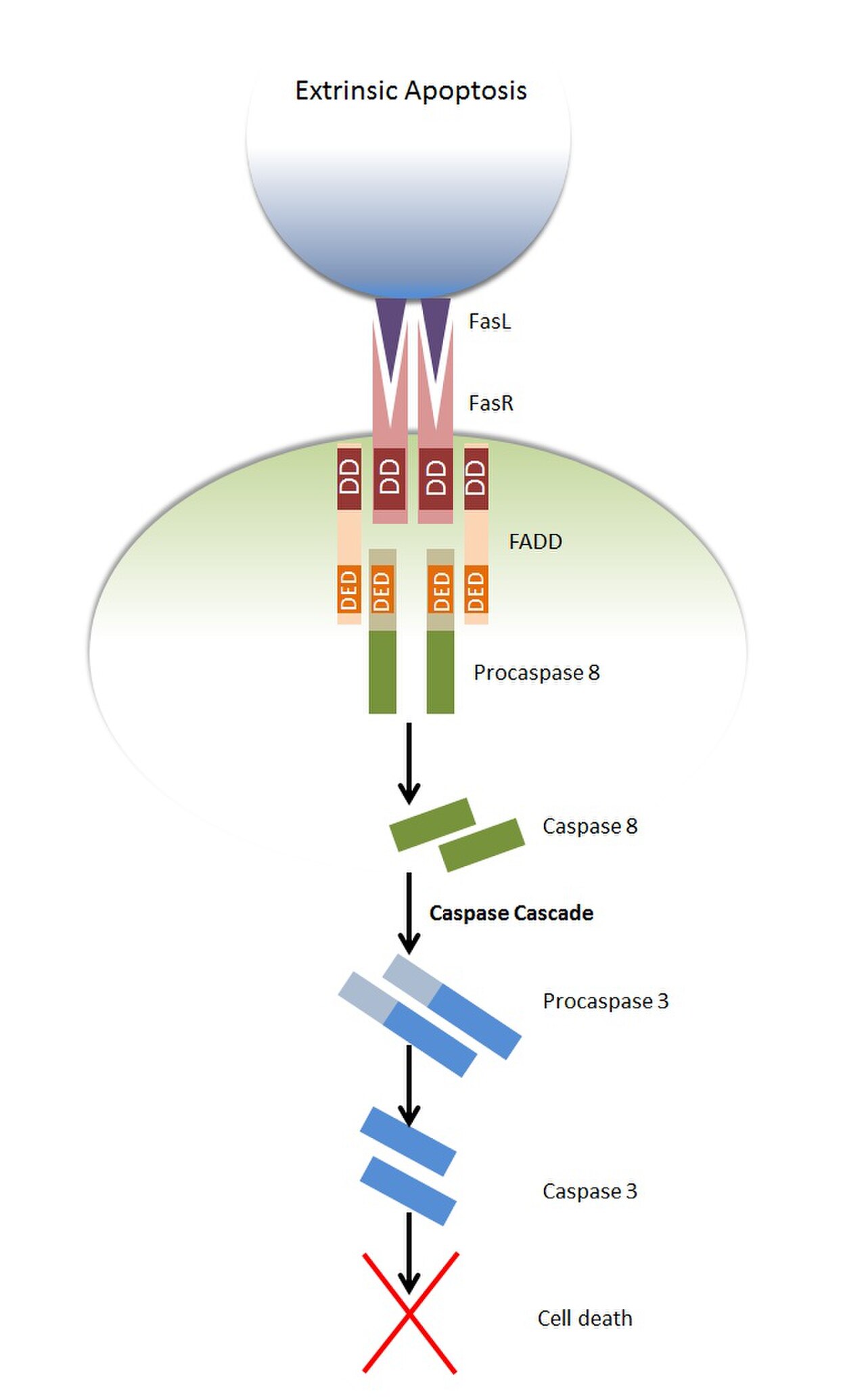
Apoptose extrinsèque
Lors de la stimulation par le ligand Fas , le récepteur Fas se trimérise. De nombreux récepteurs, dont Fas, contiennent un domaine de mort cytoplasmique et sont donc appelés récepteurs de mort . Le FADD se lie au domaine de mort de cette structure trimérique via son domaine de mort ce qui entraîne le démasquage du domaine de mort de FADD et le recrutement ultérieur des procaspases 8 et 10 via une interaction entre les domaines de mort de FADD et des procaspases. Cela génère un complexe connu sous le nom de complexe de signalisation induisant la mort (DISC). Les procaspases 8 et 10 sont connues sous le nom de caspases initiatrices . Ce sont des molécules inactives, mais lorsqu'elles sont amenées à proximité d'autres procaspases du même type, un clivage autocatalytique se produit au niveau d'un résidu d'aspartate dans leurs propres structures, ce qui donne une protéine activée. Cette protéine activée peut ensuite continuer à cliver et à activer d'autres caspases, initiant la cascade de caspases . Les caspases activées peuvent ensuite cliver des protéines intracellulaires telles que l'inhibiteur de la DNase activée par la caspase (ICAD), ce qui conduit finalement à l'apoptose de la cellule.
La liaison de TRAIL aux récepteurs de mort quatre et cinq ( DR4 et DR5 ) peut conduire à l'apoptose par le même mécanisme.
L'apoptose peut également être déclenchée par la liaison d'un ligand au récepteur 1 du facteur de nécrose tumorale (TNFR1) ; cependant, le mécanisme par lequel cela se produit est légèrement plus complexe. Une autre protéine adaptatrice contenant du DD appelée TRADD , ainsi que d'autres protéines, se lie au TNF1R activé, formant ce que l'on appelle le complexe I. Cela entraîne l'activation de la voie NFκB , qui favorise la survie cellulaire. Ce complexe est ensuite internalisé et FADD se lie à TRADD via une interaction des DD des deux protéines adaptatrices, formant ce que l'on appelle le complexe II. FADD recrute à nouveau la procaspase 8, qui initie la cascade de caspases conduisant à l'apoptose.
Nécroptose
La FADD joue également un rôle dans la régulation de la nécroptose , un processus nécessitant les kinases sérine/thréonine RIPK1 et RIPK3 . La caspase 8 activée clive ces kinases , inhibant la nécroptose. Étant donné que l'activation de la caspase 8 nécessite la FADD pour rapprocher les molécules de procaspase 8 les unes des autres afin de faciliter leur activation, la FADD est nécessaire pour réguler négativement la nécroptose. En conséquence, les cellules déficientes en FADD induisent la nécroptose car elles sont incapables de recruter et d'activer la procaspase 8. La FADD peut également se lier directement à RIPK1 et RIPK3, mais la signification de cette interaction n'est pas encore claire.
Mort cellulaire autophagique
L'autophagie est un processus qui permet la survie cellulaire dans des conditions de stress mais peut également conduire à la mort cellulaire.
Grâce à sa DD, FADD interagit avec ATG5 , une protéine impliquée dans l'autophagie. Cette interaction s'est avérée essentielle à la mort cellulaire autophagique, qui est induite par l'IFN-γ .
En revanche, il a également été démontré qu'il inhibe la mort cellulaire autophagique et favorise donc la survie cellulaire. FADD se lie à ATG5 dans un complexe qui contient également ATG12 , Caspase 8 et RIPK1. La formation de ce complexe est stimulée par la signalisation autophagique. La caspase 8 clive ensuite RIPK1, ce qui entraîne l'inhibition de cette signalisation et donc la mort cellulaire.
Développement
L'inactivation du gène FADD dans les embryons de souris est mortelle, ce qui montre que le gène FADD joue un rôle dans le développement embryonnaire . On pense que cela est dû à un développement anormal du cœur . Ce développement cardiaque anormal peut être dû à la régulation de la voie NFκB dépendante du gène FADD.
Le FADD joue également un rôle dans le développement des yeux du poisson zèbre .
Régulation du cycle cellulaire
On pense que la FADD joue un rôle dans la régulation du cycle cellulaire des lymphocytes T. Cette régulation dépend de la phosphorylation de la FADD sur la sérine 194, qui est réalisée par la caséine kinase 1a (CKIα). Cette forme phosphorylée de la FADD se trouve principalement dans le noyau et l'abondance de la FADD phosphorylée augmente significativement dans la phase G2 du cycle cellulaire par rapport à la phase G1 où on ne peut en détecter que très peu. Comme elle se trouve au niveau du fuseau mitotique pendant la phase G2, il a été proposé qu'elle intervienne dans la transition G2/M, cependant, le mécanisme par lequel elle le fait n'est pas encore connu.
Prolifération des lymphocytes
La FADD est essentielle à la prolifération des cellules T lorsque le récepteur des cellules T est stimulé par l'antigène . En revanche, la FADD n'a aucun effet sur la prolifération des cellules B induite par la stimulation du récepteur des cellules B. Cependant, elle est nécessaire à la prolifération des cellules B induite par la stimulation de TLR3 et TLR4 .
Inflammation
L'activation de la signalisation du facteur nucléaire kappa B (NFκB) conduit à la transcription de diverses cytokines pro-inflammatoires ainsi que de gènes anti-apoptotiques. Il a été constaté que la signalisation du NFκB était inhibée dans les cellules déficientes en FADD après stimulation des récepteurs TNF-R1 ou Fas. Cela suggère un rôle du FADD dans l'activation de la voie NFκB. Inversement, le FADD a également un rôle dans l'inhibition de cette voie. Normalement, lors de la stimulation des récepteurs TL4 ou IL-1R1 , la protéine adaptatrice, MyD88 , est recrutée sur la membrane plasmique où elle se lie à la kinase associée au récepteur de l'IL-1 (IRAK) via une interaction DD-DD. Cela active une voie de signalisation qui entraîne la translocation du NFκB vers le noyau, où il induit la transcription des cytokines inflammatoires. FADD peut interférer avec l'interaction entre MyD88 et IRAK, en se liant à MyD88 via son DD et donc cela perturbe la cascade qui conduirait à la translocation de NFκB et à l'inflammation.
Autre
La FADD est nécessaire pour une réponse antivirale efficace. Lors d'une infection virale, la FADD est nécessaire pour augmenter les niveaux d' Irf7, une molécule nécessaire à la production d' IFN-α . L'IFN-α est une molécule clé impliquée dans la réponse contre les virus .
La FADD est impliquée dans l'activation des phosphatases qui déphosphorylent et désactivent la protéine kinase C (PKC). Sans FADD, la PKC reste active et est capable de poursuivre les cascades de signalisation conduisant à des processus tels que les réarrangements du cytosquelette et la motilité cellulaire .
Des recherches récentes ont également montré qu’il pourrait jouer un rôle dans la régulation des niveaux de glucose et que la forme phosphorylée de FADD est importante pour cette fonction.
Règlement

Localisation subcellulaire
Le FADD peut être présent à la fois dans le noyau et dans le cytoplasme des cellules. La phosphorylation de Ser194 du FADD chez l'homme (ou Ser191 chez la souris) régulerait sa localisation subcellulaire. Une séquence de localisation nucléaire et un signal d'exportation nucléaire , tous deux situés dans le DED du FADD, sont également nécessaires pour qu'il entre et sorte du noyau. Selon sa localisation subcellulaire , le FADD peut avoir différents rôles. Dans le cytoplasme, sa principale fonction est d'induire l'apoptose. Cependant, dans le noyau, il peut avoir l'effet inverse et favoriser la survie.
c-FLIP
La protéine inhibitrice de FLICE cellulaire (c-FLIP) est une protéine régulatrice qui contient deux DED. Il existe deux isoformes de C-FLIP : C-FLIP S et FLIP L. On pensait à l'origine qu'elle agissait comme un régulateur négatif de l'apoptose en se liant au DED de FADD et en empêchant ainsi la procaspase 8 de se lier et d'inhiber la formation du DISC. Cependant, il a été observé que c-FLIP et la procaspase 8 peuvent être trouvées au niveau du même DISC. Par conséquent, il a été proposé que la présence de c-FLIP inhibe l'interaction étroite des procaspases entre elles. Sans cette proximité, les procaspases ne peuvent pas être complètement clivées et restent dans un état inactif.
PKC
L'activité de la protéine kinase C a un effet négatif sur l'apoptose médiée par le récepteur Fas. En effet, elle inhibe le recrutement de FADD vers le récepteur et donc un DISC n'est pas formé. Il a été démontré qu'en augmentant ou en diminuant la quantité de PKC dans les cellules T, plus ou moins de FADD est recruté vers FasR respectivement, lorsque le FasR est stimulé.
MKRN1
MKRN1 est une ubiquitine ligase E3 qui régule négativement la FADD en la ciblant pour la dégradation médiée par l'ubiquitine. Ce faisant, MKRN1 est capable de contrôler le niveau d'apoptose.
Rôles dans les maladies inflammatoires
Des niveaux accrus de FADD ont été trouvés dans les leucocytes de patients atteints de sclérose en plaques rémittente-récurrente , contribuant à l'inflammation . Dans la polyarthrite rhumatoïde , on pense que la stimulation des récepteurs Fas sur les macrophages conduit à la formation de FADD contenant des DISC. La formation de ces séquestrants éloigne FADD de MyD88, ce qui permet à MyD88 d'interagir avec IRAK et d'induire l'inflammation accrue associée à cette maladie.
Rôles dans le cancer
Comme FADD joue un rôle si important dans l'apoptose, la perte de FADD peut donner aux cellules cancéreuses un avantage prolifératif puisque l'apoptose ne serait plus induite lorsque les récepteurs Fas sont stimulés.
Cependant, il existe une régulation positive significative de FADD dans le cancer de l'ovaire et le carcinome épidermoïde de la tête et du cou . On ne sait pas encore clairement quel avantage cela a sur les cellules cancéreuses, mais étant donné le rôle des FADD dans la régulation du cycle cellulaire et la survie cellulaire, il est probable que cela puisse être lié à cela. On observe également des niveaux élevés de FADD dans le cancer du poumon non à petites cellules . FADD peut être utilisé comme marqueur pronostique pour ces deux maladies, des niveaux élevés de FADD étant corrélés à un mauvais pronostic.
Cible thérapeutique
Le taxol est un médicament utilisé dans les thérapies anticancéreuses en raison de sa capacité à interférer avec l'assemblage des microtubules , ce qui conduit à l'arrêt du cycle cellulaire. Le FADD phosphorylé au niveau de Ser194 rend les cellules plus sensibles à l'arrêt du cycle cellulaire induit par le taxol. Le taxol peut également provoquer l'apoptose des cellules et cela nécessite la procaspase 10, qui est activée par le recrutement au FADD.
Il a été démontré que l'activation de JNK conduit à la phosphorylation de FADD. FADD phosphorylé peut induire un arrêt du cycle cellulaire G2/M, potentiellement en augmentant la stabilité de p53. Par conséquent, les médicaments qui peuvent activer cette voie peuvent avoir un potentiel thérapeutique. Cependant, des niveaux élevés de FADD phosphorylé ont été corrélés à un mauvais pronostic dans de nombreux cancers tels que celui de la tête et du cou. Cela est probablement dû à son activation de la voie NF-κB, qui est anti-apoptotique. Par conséquent, l'inhibition de la phosphorylation de FADD peut être développée comme une stratégie potentielle de lutte contre le cancer. Par exemple, il a été suggéré que l'inhibition de FADD pourrait fonctionner comme une thérapie ciblée potentielle pour le cancer de l'ovaire résistant aux médicaments .
Interactions
Il a été observé que FADD interagissait avec le récepteur Fas :
- ABCA1 ,
- ATG5 ,
- C-FLIP , MKRN1,
- Caséine kinase 1a ,
- DEDD ,
- MBD4
- MonD88 ,
- NACA ,
- PEA15 ,
- RIPK1 ,
- RIPK3 ,
- TRAD ,
- SENTIER ,
- procaspase 10 , et
- Procaspase 8 .